

Specific glycine-dependent enzyme motion determines the potency of conformation selective inhibitors of threonyl-tRNA synthetase
- PMID: 39014102
- PMCID: PMC11252418
- DOI: 10.1038/s42003-024-06559-x
Specific glycine-dependent enzyme motion determines the potency of conformation selective inhibitors of threonyl-tRNA synthetase
Abstract
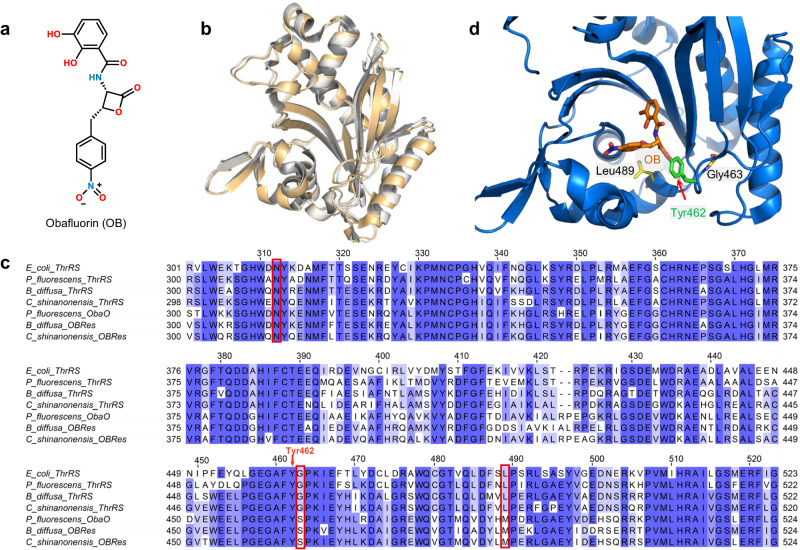
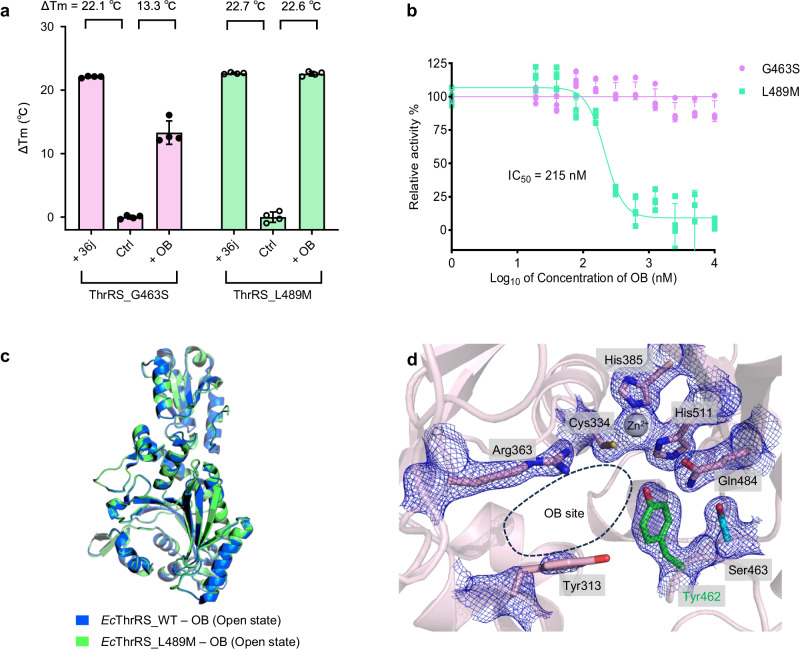
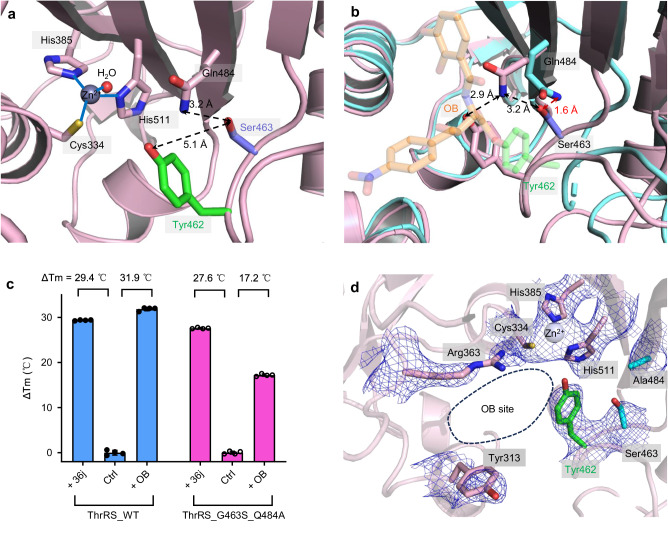
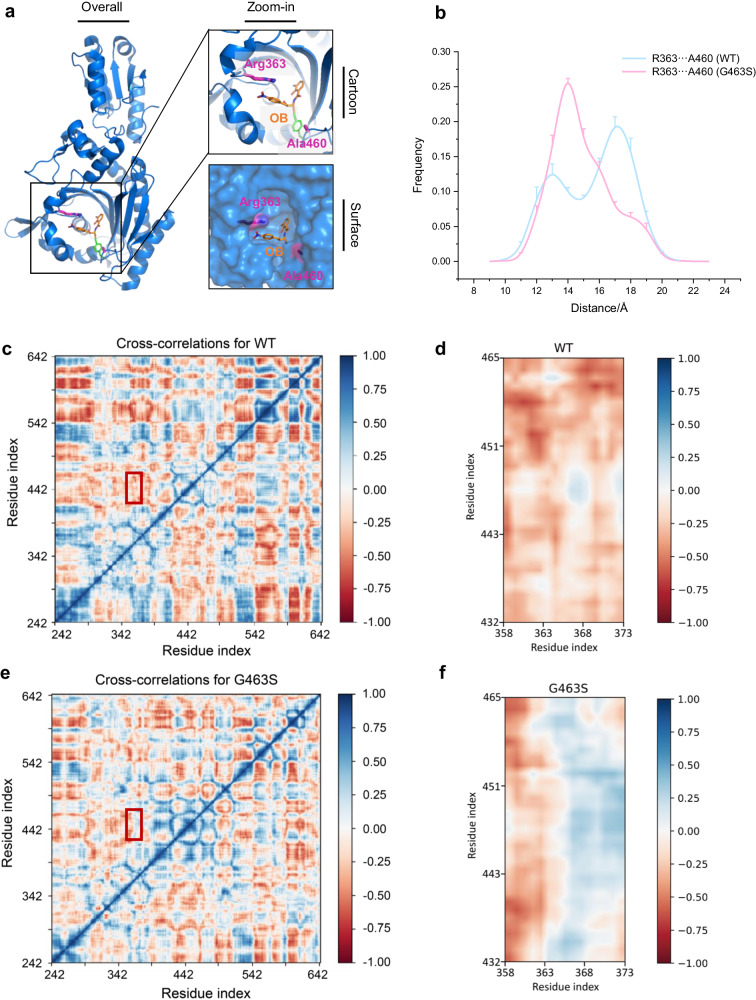
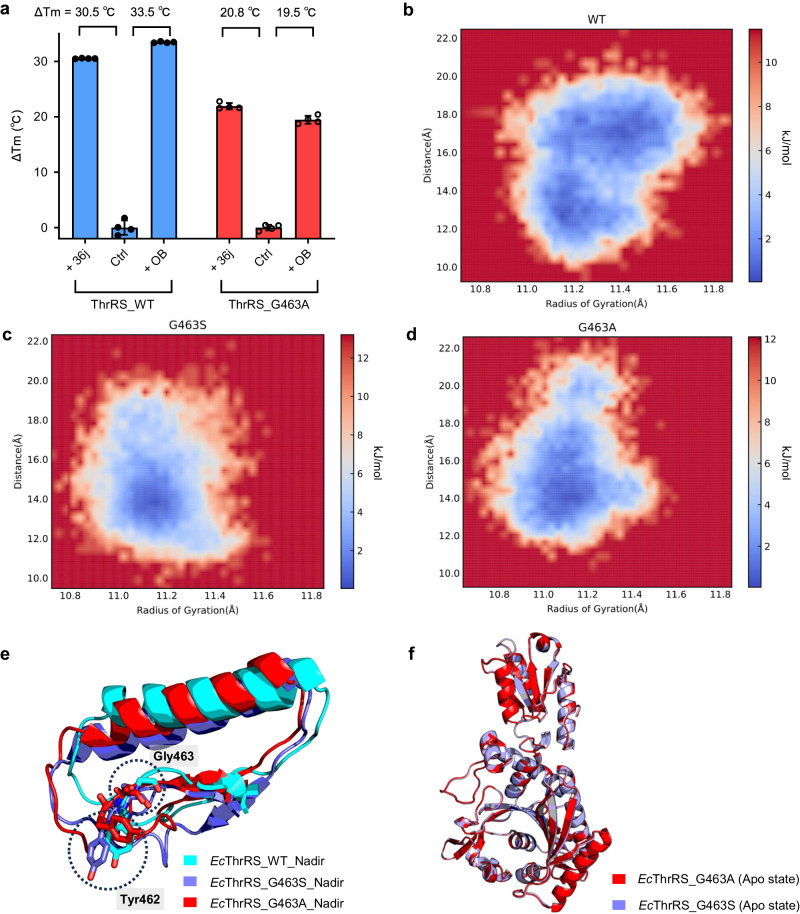
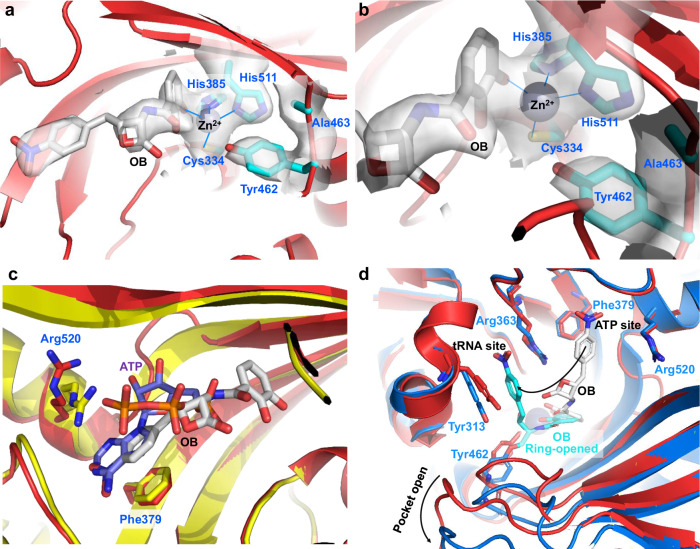
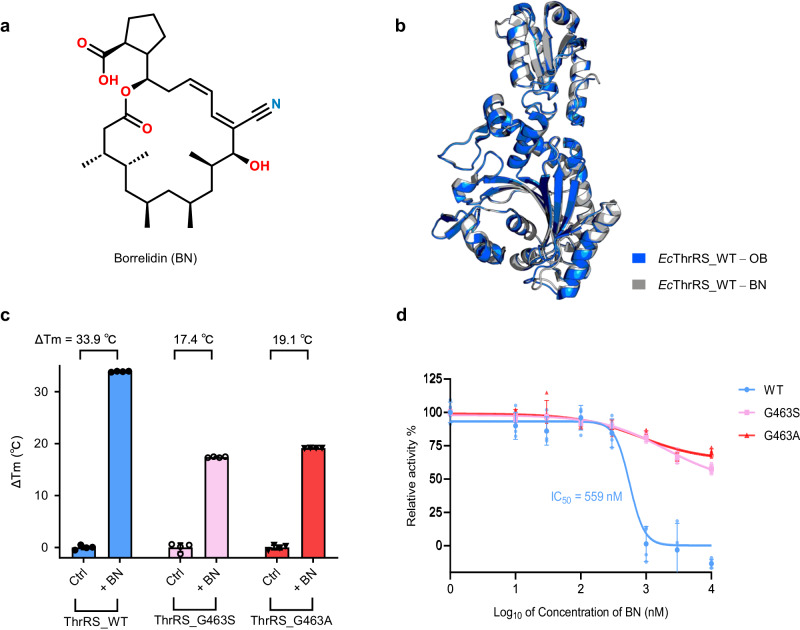
The function of proteins depends on their correct structure and proper dynamics. Understanding the dynamics of target proteins facilitates drug design and development. However, dynamic information is often hidden in the spatial structure of proteins. It is important but difficult to identify the specific residues that play a decisive role in protein dynamics. Here, we report that a critical glycine residue (Gly463) dominates the motion of threonyl-tRNA synthetase (ThrRS) and the sensitivity of the enzyme to antibiotics. Obafluorin (OB), a natural antibiotic, is a novel covalent inhibitor of ThrRS. The binding of OB induces a large conformational change in ThrRS. Through five crystal structures, biochemical and biophysical analyses, and computational simulations, we found that Gly463 plays an important role in the dynamics of ThrRS. Mutating this flexible residue into more rigid residues did not damage the enzyme's three-dimensional structure but significantly improved the thermal stability of the enzyme and suppressed its ability to change conformation. These mutations cause resistance of ThrRS to antibiotics that are conformationally selective, such as OB and borrelidin. This work not only elucidates the molecular mechanism of the self-resistance of OB-producing Pseudomonas fluorescens but also emphasizes the importance of backbone kinetics for aminoacyl-tRNA synthetase-targeting drug development.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
-
Identification of a novel toxicophore in anti-cancer chemotherapeutics that targets mitochondrial respiratory complex I.Elife. 2020 May 20;9:e55845. doi: 10.7554/eLife.55845. Elife. 2020. PMID: 32432547 Free PMC article.
-
Far Posterior Approach for Rib Fracture Fixation: Surgical Technique and Tips.JBJS Essent Surg Tech. 2024 Dec 6;14(4):e23.00094. doi: 10.2106/JBJS.ST.23.00094. eCollection 2024 Oct-Dec. JBJS Essent Surg Tech. 2024. PMID: 39650795 Free PMC article.
-
Screening a library of potential competitive inhibitors against bacterial threonyl-tRNA synthetase: DFT calculations.J Biomol Struct Dyn. 2024;42(24):13555-13563. doi: 10.1080/07391102.2023.2276878. Epub 2023 Nov 1. J Biomol Struct Dyn. 2024. PMID: 37909495
-
Conservative, physical and surgical interventions for managing faecal incontinence and constipation in adults with central neurological diseases.Cochrane Database Syst Rev. 2024 Oct 29;10(10):CD002115. doi: 10.1002/14651858.CD002115.pub6. Cochrane Database Syst Rev. 2024. PMID: 39470206
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
