

The root-knot nematode effector MiEFF12 targets the host ER quality control system to suppress immune responses and allow parasitism
- PMID: 38961768
- PMCID: PMC11222708
- DOI: 10.1111/mpp.13491
The root-knot nematode effector MiEFF12 targets the host ER quality control system to suppress immune responses and allow parasitism
Abstract
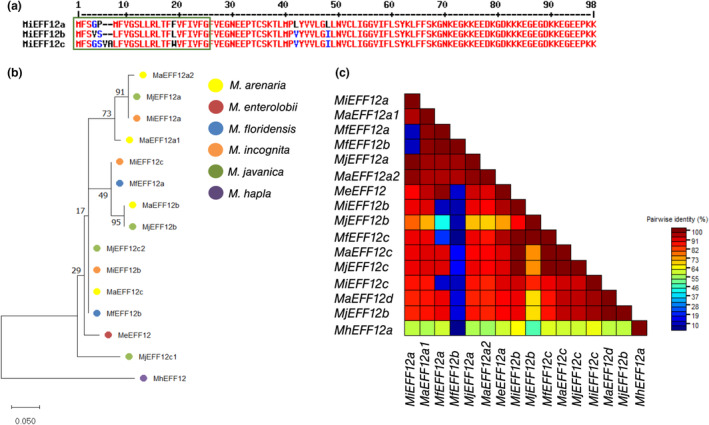
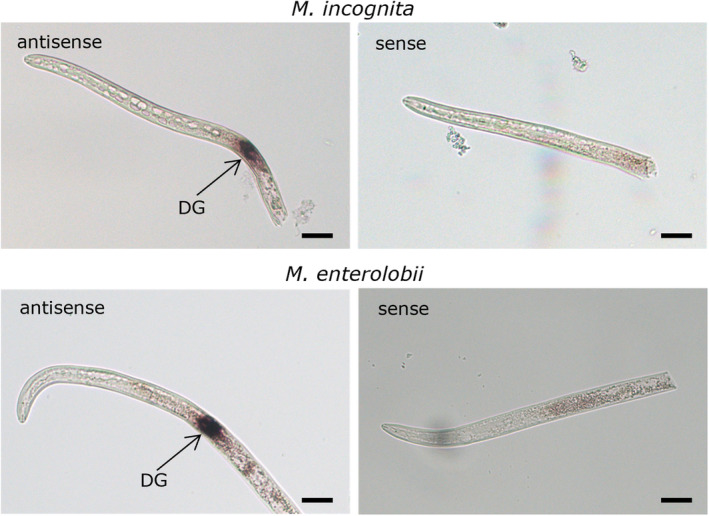
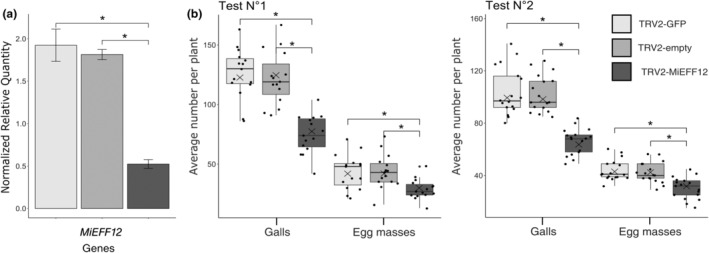
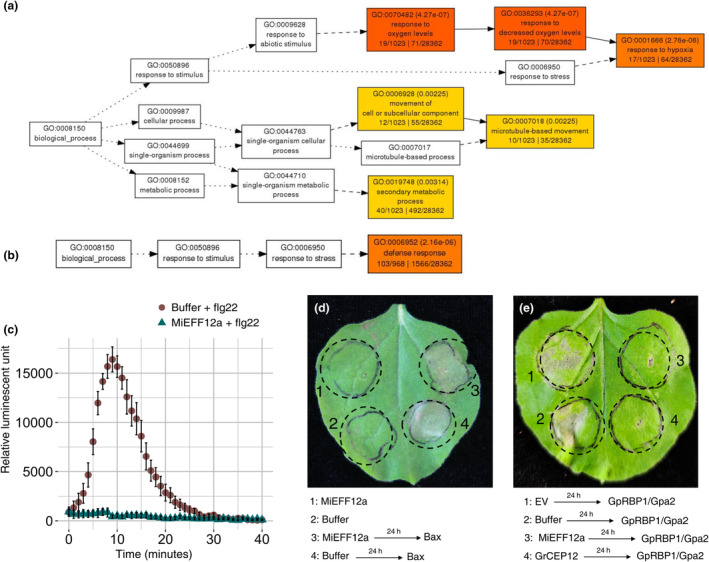
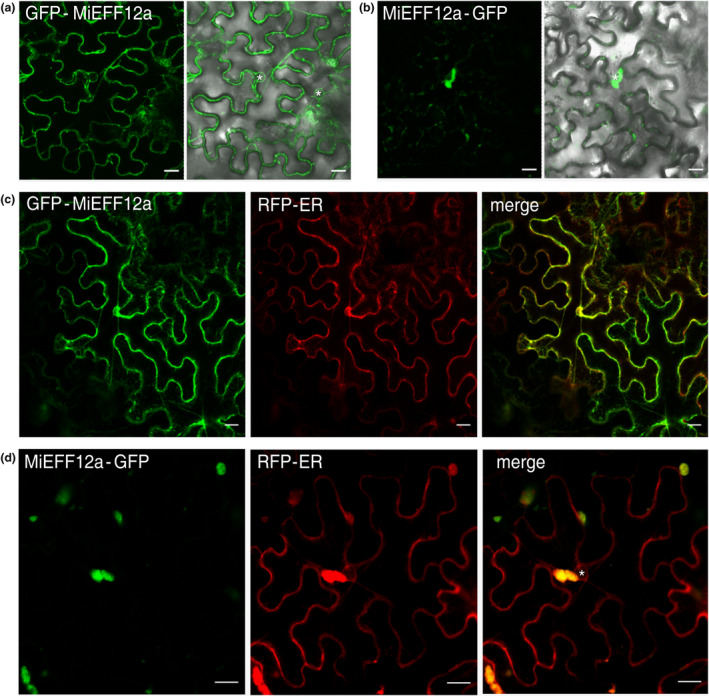
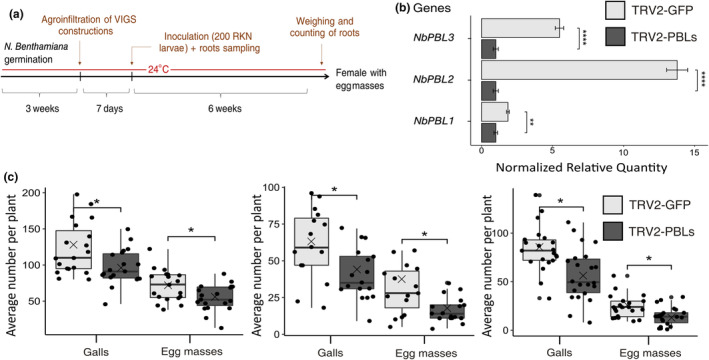
Root-knot nematodes (RKNs) are microscopic parasitic worms able to infest the roots of thousands of plant species, causing massive crop yield losses worldwide. They evade the plant's immune system and manipulate plant cell physiology and metabolism to transform a few root cells into giant cells, which serve as feeding sites for the nematode. RKN parasitism is facilitated by the secretion in planta of effector molecules, mostly proteins that hijack host cellular processes. We describe here a conserved RKN-specific effector, effector 12 (EFF12), that is synthesized exclusively in the oesophageal glands of the nematode, and we demonstrate its function in parasitism. In the plant, MiEFF12 localizes to the endoplasmic reticulum (ER). A combination of RNA-sequencing analysis and immunity-suppression bioassays revealed the contribution of MiEFF12 to the modulation of host immunity. Yeast two-hybrid, split luciferase and co-immunoprecipitation approaches identified an essential component of the ER quality control system, the Solanum lycopersicum plant bap-like (PBL), and basic leucine zipper 60 (BZIP60) proteins as host targets of MiEFF12. Finally, silencing the PBL genes in Nicotiana benthamiana decreased susceptibility to Meloidogyne incognita infection. Our results suggest that EFF12 manipulates PBL function to modify plant immune responses to allow parasitism.
Keywords: Meloidogyne incognita; Nicotiana benthamiana; Solanum lycopersicum; ER quality control; effector; endoplasmic reticulum (ER).
© 2024 The Author(s). Molecular Plant Pathology published by British Society for Plant Pathology and John Wiley & Sons Ltd.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures

Similar articles
-
A root-knot nematode effector targets the Arabidopsis cysteine protease RD21A for degradation to suppress plant defense and promote parasitism.Plant J. 2024 Jun;118(5):1500-1515. doi: 10.1111/tpj.16692. Epub 2024 Mar 22. Plant J. 2024. PMID: 38516730
-
Root-knot nematodes exploit the catalase-like effector to manipulate plant reactive oxygen species levels by directly degrading H2O2.Mol Plant Pathol. 2024 Sep;25(9):e70000. doi: 10.1111/mpp.70000. Mol Plant Pathol. 2024. PMID: 39254175 Free PMC article.
-
Rice susceptibility to root-knot nematodes is enhanced by the Meloidogyne incognita MSP18 effector gene.Planta. 2019 Oct;250(4):1215-1227. doi: 10.1007/s00425-019-03205-3. Epub 2019 Jun 19. Planta. 2019. PMID: 31218413
-
Manipulation of plant cells by cyst and root-knot nematode effectors.Mol Plant Microbe Interact. 2013 Jan;26(1):9-16. doi: 10.1094/MPMI-05-12-0106-FI. Mol Plant Microbe Interact. 2013. PMID: 22809272 Review.
-
Manipulation of host gene expression by root-knot nematodes.J Parasitol. 1996 Dec;82(6):881-8. J Parasitol. 1996. PMID: 8973394 Review.
References
-
- Abad, P. , Gouzy, J. , Aury, J.M. , Castagnone‐Sereno, P. , Danchin, E.G. , Deleury, E. et al. (2008) Genome sequence of the metazoan plant‐parasitic nematode Meloidogyne incognita . Nature Biotechnology, 26, 909–915. - PubMed
-
- Atabekova, A.K. , Pankratenko, A.V. , Makarova, S.S. , Lazareva, E.A. , Owens, R.A. , Solovyev, A.G. et al. (2017) Phylogenetic and functional analyses of a plant protein related to human B‐cell receptor‐associated proteins. Biochimie, 132, 28–37. - PubMed
-
- Berger, A. , Boscari, A. , Horta Araújo, N. , Maucourt, M. , Hanchi, M. , Bernillon, S. et al. (2020) Plant nitrate reductases regulate nitric oxide production and nitrogen‐fixing metabolism during the Medicago truncatula–Sinorhizobium meliloti symbiosis. Frontiers in Plant Science, 11, 1313. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
