

DNMT1 can induce primary germ layer differentiation through de novo DNA methylation
- PMID: 38811355
- PMCID: PMC11447926
- DOI: 10.1111/gtc.13130
DNMT1 can induce primary germ layer differentiation through de novo DNA methylation
Abstract
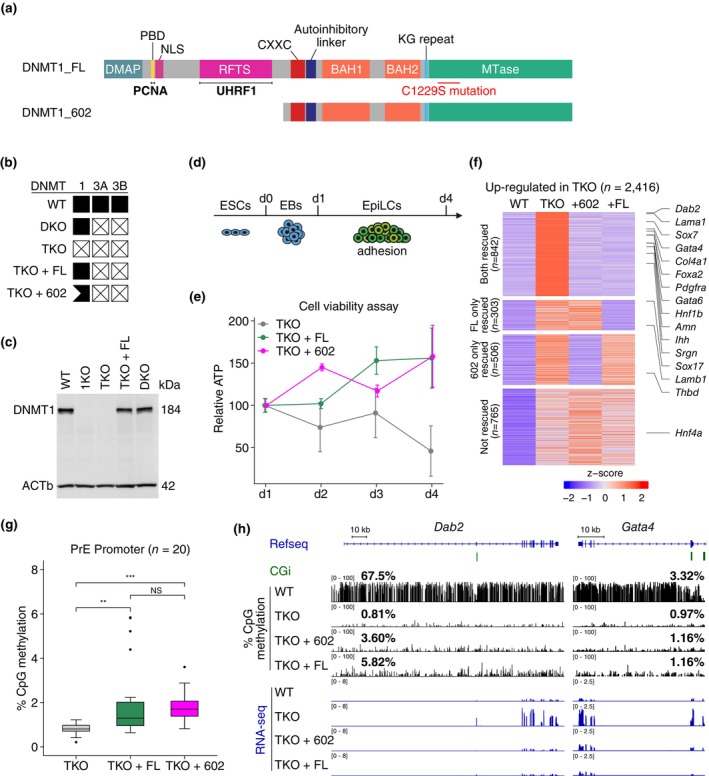
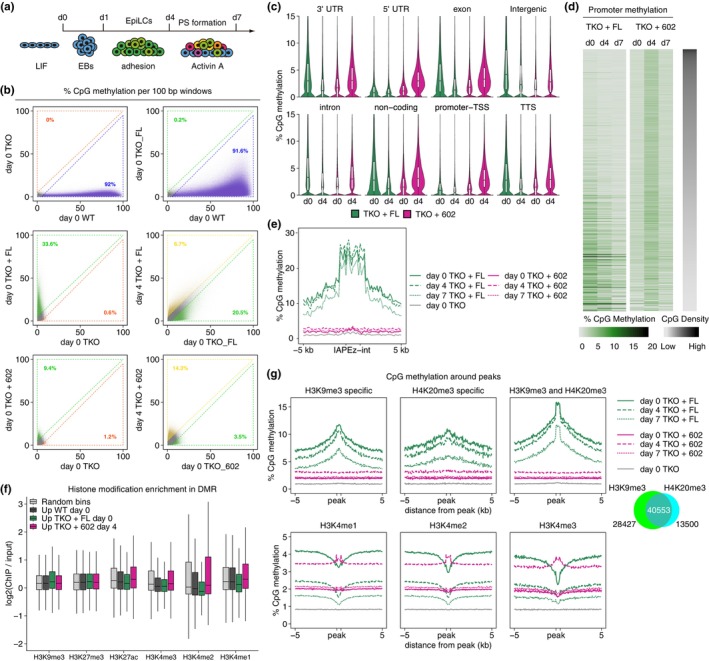
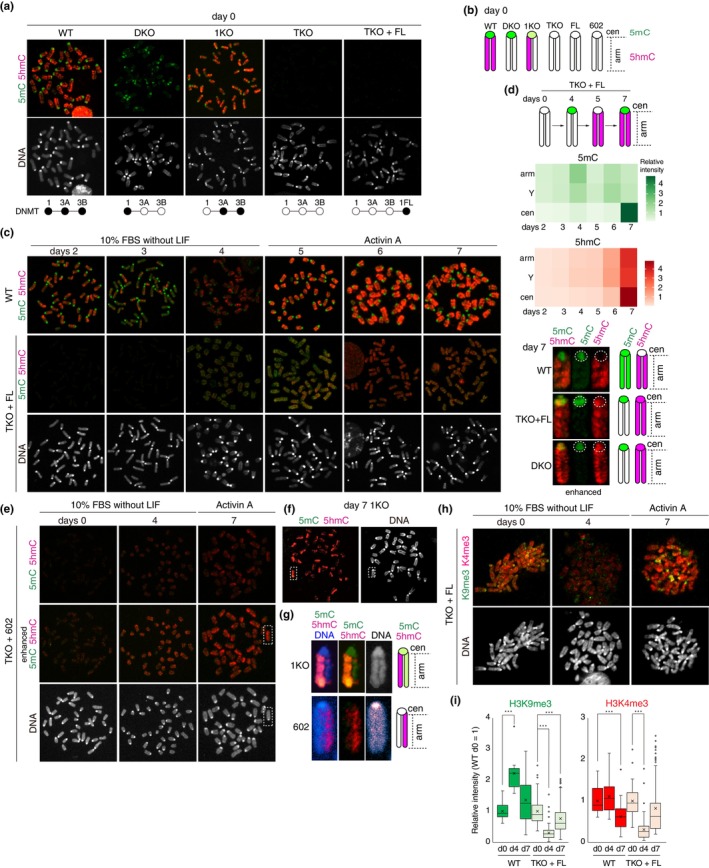
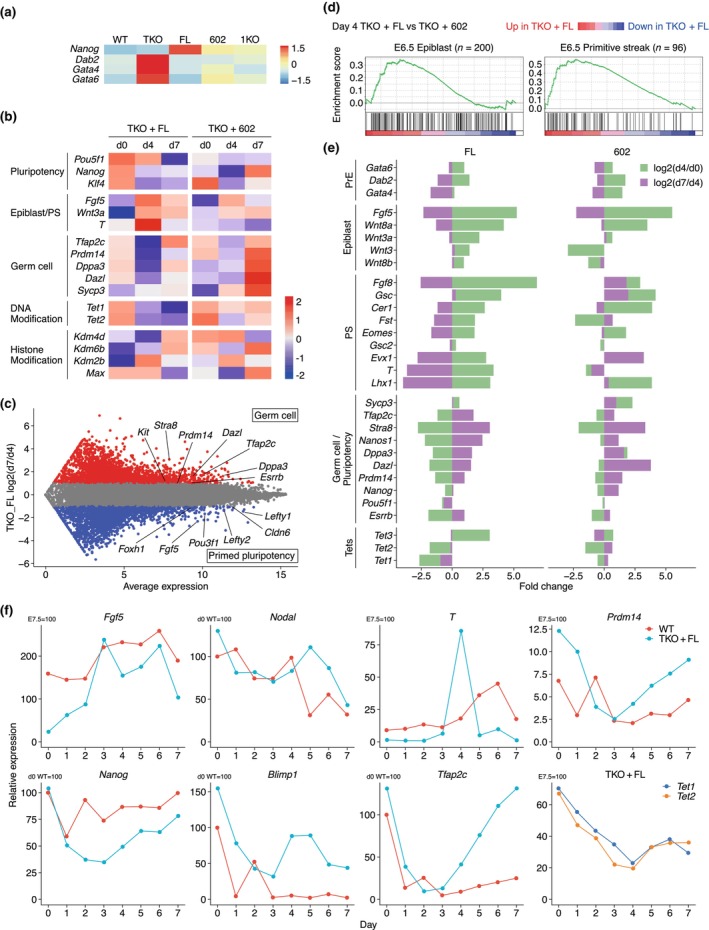
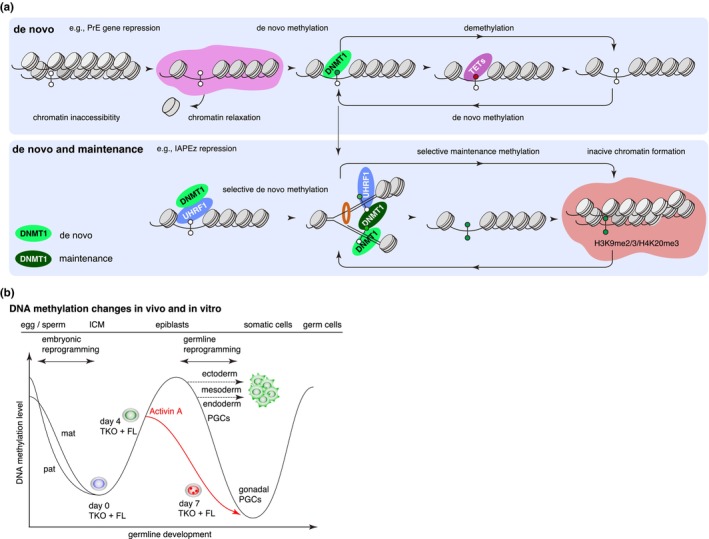
DNA methyltransferases and Ten-Eleven Translocation (TET) proteins regulate the DNA methylation and demethylation cycles during mouse embryonic development. Although DNMT1 mainly plays a role in the maintenance of DNA methylation after DNA replication, it is also reported to possess de novo methyltransferase capacity. However, its physiological significance remains unclear. Here, we demonstrate that full-length DNMT1 (FL) and a mutant lacking the N-terminus necessary for its maintenance activity (602) confer the differentiation potential of mouse Dnmt1, Dnmt3a, and Dnmt3b (Dnmts-TKO) embryonic stem cells (ESCs). Both FL and 602 inhibit the spontaneous differentiation of Dnmts-TKO ESCs in the undifferentiated state. Dnmts-TKO ESCs showed loss of DNA methylation and de-repression of primitive endoderm-related genes, but these defects were partially restored in Dnmts-TKO + FL and Dnmts-TKO + 602 ESCs. Upon differentiation, Dnmts-TKO + FL ESCs show increased 5mC and 5hmC levels across chromosomes, including pericentromeric regions. In contrast, Dnmts-TKO + 602 ESCs didn't accumulate 5mC, and sister chromatids showed 5hmC asynchronously. Furthermore, in comparison with DNMT1_602, DNMT1_FL effectively promoted commitment to the epiblast-like cells and beyond, driving cell-autonomous mesendodermal and germline differentiation through embryoid body-based methods. With precise target selectivity achieved by its N-terminal region, DNMT1 may play a role in gene regulation leading to germline development.
Keywords: DNA methylation; Dnmt1; embryonic stem cells; epigenetics.
© 2024 The Author(s). Genes to Cells published by Molecular Biology Society of Japan and John Wiley & Sons Australia, Ltd.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
DNMT1 regulates the timing of DNA methylation by DNMT3 in an enzymatic activity-dependent manner in mouse embryonic stem cells.PLoS One. 2022 Jan 5;17(1):e0262277. doi: 10.1371/journal.pone.0262277. eCollection 2022. PLoS One. 2022. PMID: 34986190 Free PMC article.
-
Global DNA hypomethylation prevents consolidation of differentiation programs and allows reversion to the embryonic stem cell state.PLoS One. 2012;7(12):e52629. doi: 10.1371/journal.pone.0052629. Epub 2012 Dec 27. PLoS One. 2012. PMID: 23300728 Free PMC article.
-
Widespread recovery of methylation at gametic imprints in hypomethylated mouse stem cells following rescue with DNMT3A2.Epigenetics Chromatin. 2016 Nov 22;9:53. doi: 10.1186/s13072-016-0104-2. eCollection 2016. Epigenetics Chromatin. 2016. PMID: 27895716 Free PMC article.
-
DNA methyltransferases in hematological malignancies.J Genet Genomics. 2020 Jul 20;47(7):361-372. doi: 10.1016/j.jgg.2020.04.006. Epub 2020 Jul 24. J Genet Genomics. 2020. PMID: 32994141 Free PMC article. Review.
-
Role of Mammalian DNA Methyltransferases in Development.Annu Rev Biochem. 2020 Jun 20;89:135-158. doi: 10.1146/annurev-biochem-103019-102815. Epub 2019 Dec 9. Annu Rev Biochem. 2020. PMID: 31815535 Review.
References
-
- Aramaki, S. , Hayashi, K. , Kurimoto, K. , Ohta, H. , Yabuta, Y. , Iwanari, H. , Mochizuki, Y. , Hamakubo, T. , Kato, Y. , Shirahige, K. , & Saitou, M. (2013). A mesodermal factor, T, specifies mouse germ cell fate by directly activating germline determinants. Developmental Cell, 27, 516–529. - PubMed
-
- Barau, J. , Teissandier, A. , Zamudio, N. , Roy, S. , Nalesso, V. , Hérault, Y. , Guillou, F. , & Bourc'his, D. (2016). The DNA methyltransferase DNMT3C protects male germ cells from transposon activity. Science, 354, 912. - PubMed
-
- Beck, S. , Le Good, J. A. , Guzman, M. , Haim, N. B. , Roy, K. , Beermann, F. , & Constam, D. B. (2002). Extraembryonic proteases regulate nodal signalling during gastrulation. Nature Cell Biology, 4, 981–985. - PubMed
-
- Becker, S. , Casanova, J. , & Grabel, L. (1992). Localization of endoderm‐specific mRNAs in differentiating F9 embryoid bodies. Mechanisms of Development, 37, 3–12. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
