

Connexin Gap Junction Channels and Hemichannels: Insights from High-Resolution Structures
- PMID: 38785780
- PMCID: PMC11117596
- DOI: 10.3390/biology13050298
Connexin Gap Junction Channels and Hemichannels: Insights from High-Resolution Structures
Abstract
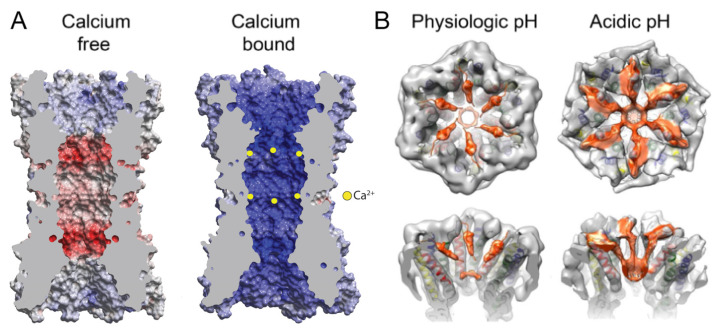
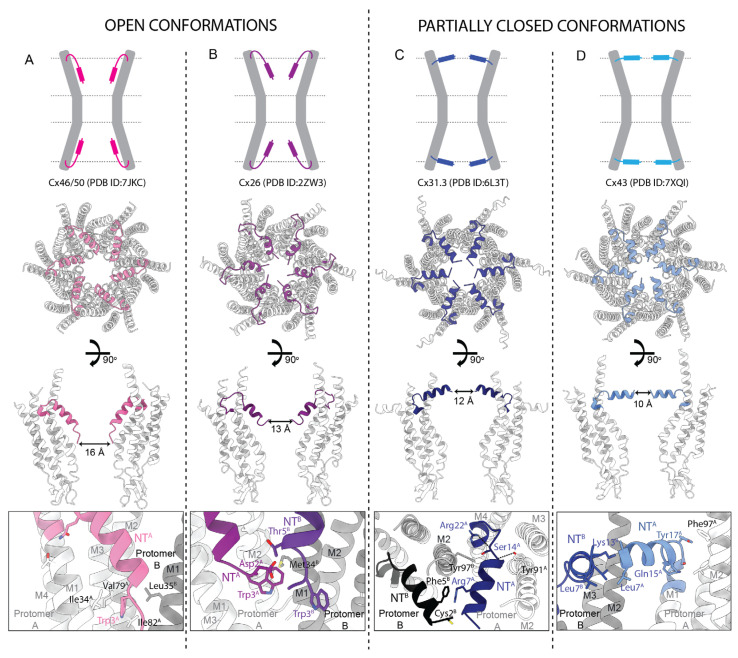
Connexins (Cxs) are a family of integral membrane proteins, which function as both hexameric hemichannels (HCs) and dodecameric gap junction channels (GJCs), behaving as conduits for the electrical and molecular communication between cells and between cells and the extracellular environment, respectively. Their proper functioning is crucial for many processes, including development, physiology, and response to disease and trauma. Abnormal GJC and HC communication can lead to numerous pathological states including inflammation, skin diseases, deafness, nervous system disorders, and cardiac arrhythmias. Over the last 15 years, high-resolution X-ray and electron cryomicroscopy (cryoEM) structures for seven Cx isoforms have revealed conservation in the four-helix transmembrane (TM) bundle of each subunit; an αβ fold in the disulfide-bonded extracellular loops and inter-subunit hydrogen bonding across the extracellular gap that mediates end-to-end docking to form a tight seal between hexamers in the GJC. Tissue injury is associated with cellular Ca2+ overload. Surprisingly, the binding of 12 Ca2+ ions in the Cx26 GJC results in a novel electrostatic gating mechanism that blocks cation permeation. In contrast, acidic pH during tissue injury elicits association of the N-terminal (NT) domains that sterically blocks the pore in a "ball-and-chain" fashion. The NT domains under physiologic conditions display multiple conformational states, stabilized by protein-protein and protein-lipid interactions, which may relate to gating mechanisms. The cryoEM maps also revealed putative lipid densities within the pore, intercalated among transmembrane α-helices and between protomers, the functions of which are unknown. For the future, time-resolved cryoEM of isolated Cx channels as well as cryotomography of GJCs and HCs in cells and tissues will yield a deeper insight into the mechanisms for channel regulation. The cytoplasmic loop (CL) and C-terminal (CT) domains are divergent in sequence and length, are likely involved in channel regulation, but are not visualized in the high-resolution X-ray and cryoEM maps presumably due to conformational flexibility. We expect that the integrated use of synergistic physicochemical, spectroscopic, biophysical, and computational methods will reveal conformational dynamics relevant to functional states. We anticipate that such a wealth of results under different pathologic conditions will accelerate drug discovery related to Cx channel modulation.
Keywords: X-ray crystallography; calcium regulation; channel gating; connexin; electron cryomicroscopy; gap junction channel; gap junction hemichannel; lipid binding; pH regulation.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures


Similar articles
-
Emerging issues of connexin channels: biophysics fills the gap.Q Rev Biophys. 2001 Aug;34(3):325-472. doi: 10.1017/s0033583501003705. Q Rev Biophys. 2001. PMID: 11838236 Review.
-
Cryo-EM structure of an open conformation of a gap junction hemichannel in lipid bilayer nanodiscs.Structure. 2021 Sep 2;29(9):1040-1047.e3. doi: 10.1016/j.str.2021.05.010. Epub 2021 Jun 14. Structure. 2021. PMID: 34129834 Free PMC article.
-
A Steric "Ball-and-Chain" Mechanism for pH-Mediated Regulation of Gap Junction Channels.Cell Rep. 2020 Apr 21;31(3):107482. doi: 10.1016/j.celrep.2020.03.046. Cell Rep. 2020. PMID: 32320665 Free PMC article.
-
Structure of the connexin-43 gap junction channel in a putative closed state.Elife. 2023 Aug 3;12:RP87616. doi: 10.7554/eLife.87616. Elife. 2023. PMID: 37535063 Free PMC article.
-
Regulation of connexin hemichannel activity by membrane potential and the extracellular calcium in health and disease.Neuropharmacology. 2013 Dec;75:479-90. doi: 10.1016/j.neuropharm.2013.03.040. Epub 2013 Apr 12. Neuropharmacology. 2013. PMID: 23587648 Review.
Cited by
-
Calcium Role in Gap Junction Channel Gating: Direct Electrostatic or Calmodulin-Mediated?Int J Mol Sci. 2024 Sep 10;25(18):9789. doi: 10.3390/ijms25189789. Int J Mol Sci. 2024. PMID: 39337278 Free PMC article. Review.
-
Calcium Regulation of Connexin Hemichannels.Int J Mol Sci. 2024 Jun 15;25(12):6594. doi: 10.3390/ijms25126594. Int J Mol Sci. 2024. PMID: 38928300 Free PMC article. Review.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
