

The haemagglutinin-neuraminidase protein of velogenic Newcastle disease virus enhances viral infection through NF-κB-mediated programmed cell death
- PMID: 38715081
- PMCID: PMC11077864
- DOI: 10.1186/s13567-024-01312-y
The haemagglutinin-neuraminidase protein of velogenic Newcastle disease virus enhances viral infection through NF-κB-mediated programmed cell death
Abstract
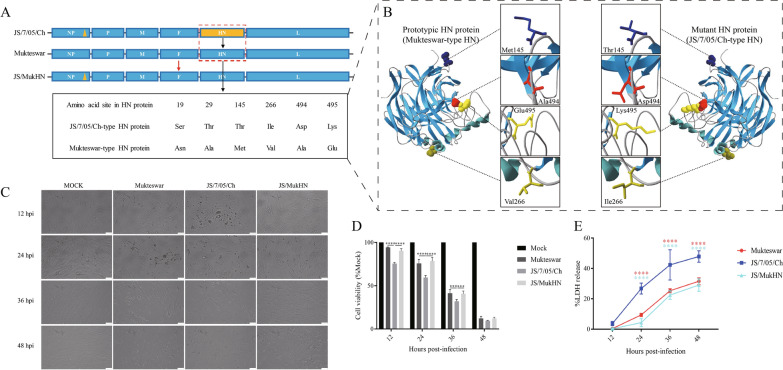
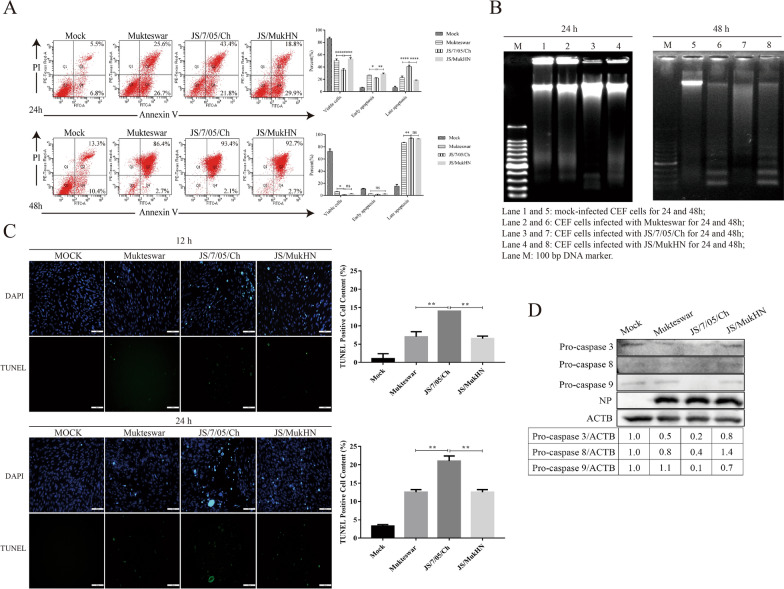
The haemagglutinin-neuraminidase (HN) protein, a vital membrane glycoprotein, plays a pivotal role in the pathogenesis of Newcastle disease virus (NDV). Previously, we demonstrated that a mutation in the HN protein is essential for the enhanced virulence of JS/7/05/Ch, a velogenic variant NDV strain originating from the mesogenic vaccine strain Mukteswar. Here, we explored the effects of the HN protein during viral infection in vitro using three viruses: JS/7/05/Ch, Mukteswar, and an HN-replacement chimeric NDV, JS/MukHN. Through microscopic observation, CCK-8, and LDH release assays, we demonstrated that compared with Mukteswar and JS/MukHN, JS/7/05/Ch intensified the cellular damage and mortality attributed to the mutant HN protein. Furthermore, JS/7/05/Ch induced greater levels of apoptosis, as evidenced by the activation of caspase-3/8/9. Moreover, JS/7/05/Ch promoted autophagy, leading to increased autophagosome formation and autophagic flux. Subsequent pharmacological experiments revealed that inhibition of apoptosis and autophagy significantly impacted virus replication and cell viability in the JS/7/05/Ch-infected group, whereas less significant effects were observed in the other two infected groups. Notably, the mutant HN protein enhanced JS/7/05/Ch-induced apoptosis and autophagy by suppressing NF-κB activation, while it mitigated the effects of NF-κB on NDV infection. Overall, our study offers novel insights into the mechanisms underlying the increased virulence of NDV and serves as a reference for the development of vaccines.
Keywords: NF-κB; Newcastle disease virus; haemagglutinin–neuraminidase; programmed cell death; viral infection.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
[Full-length genome analysis of two genotype III velogenic Newcastle diseases virus strains reveals their close relationship with vaccine Mukteswar].Wei Sheng Wu Xue Bao. 2009 Mar;49(3):302-8. Wei Sheng Wu Xue Bao. 2009. PMID: 19623952 Chinese.
-
Cellular vimentin regulates the infectivity of Newcastle disease virus through targeting of the HN protein.Vet Res. 2023 Oct 17;54(1):92. doi: 10.1186/s13567-023-01230-5. Vet Res. 2023. PMID: 37848995 Free PMC article.
-
Biological Significance of Dual Mutations A494D and E495K of the Genotype III Newcastle Disease Virus Hemagglutinin-Neuraminidase In Vitro and In Vivo.Viruses. 2022 Oct 25;14(11):2338. doi: 10.3390/v14112338. Viruses. 2022. PMID: 36366435 Free PMC article.
-
Effects of the HN Antigenic Difference between the Vaccine Strain and the Challenge Strain of Newcastle Disease Virus on Virus Shedding and Transmission.Viruses. 2017 Aug 15;9(8):225. doi: 10.3390/v9080225. Viruses. 2017. PMID: 28809802 Free PMC article.
-
Molecular Mechanisms of Anti-Neoplastic and Immune Stimulatory Properties of Oncolytic Newcastle Disease Virus.Biomedicines. 2022 Feb 28;10(3):562. doi: 10.3390/biomedicines10030562. Biomedicines. 2022. PMID: 35327364 Free PMC article. Review.
References
-
- Yan C, Liu H, Jia Y, Prince-Theodore DW, Yang M, Addoma Adam FE, Ren J, Cao X, Wang X, Xiao S, Zhang S, Yang Z. Screening and mechanistic study of key sites of the hemagglutinin-neuraminidase protein related to the virulence of Newcastle disease virus. Poult Sci. 2020;99:3374–3384. doi: 10.1016/j.psj.2020.04.014. - DOI - PMC - PubMed
-
- Amarasinghe GK, Ayllon MA, Bao Y, Basler CF, Bavari S, Blasdell KR, Briese T, Brown PA, Bukreyev A, Balkema-Buschmann A, Buchholz UJ, Chabi-Jesus C, Chandran K, Chiapponi C, Crozier I, de Swart RL, Dietzgen RG, Dolnik O, Drexler JF, Durrwald R, Dundon WG, Duprex WP, Dye JM, Easton AJ, Fooks AR, Formenty PBH, Fouchier RAM, Freitas-Astua J, Griffiths A, Hewson R, et al. Taxonomy of the order Mononegavirales: update 2019. Arch Virol. 2019;164:1967–1980. doi: 10.1007/s00705-019-04247-4. - DOI - PMC - PubMed
-
- Xu X, Qian J, Qin L, Li J, Xue C, Ding J, Wang W, Ding W, Yin R, Jin N, Ding Z. Chimeric Newcastle disease virus-like particles containing DC-binding peptide-fused haemagglutinin protect chickens from virulent Newcastle disease virus and H9N2 Avian influenza virus challenge. Virol Sin. 2020;35:455–467. doi: 10.1007/s12250-020-00199-1. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- 32302840/National Natural Science Foundation of China
- 31873021/National Natural Science Foundation of China
- 32202767/National Natural Science Foundation of China
- CARS-40/Earmarked Fund for China Agriculture Research System
- 111 Project D18007/Priority Academic Program Development of Jiangsu Higher Education Institutions
LinkOut - more resources
Full Text Sources
Research Materials
