

TIM-3+ CD8 T cells with a terminally exhausted phenotype retain functional capacity in hematological malignancies
- PMID: 38640253
- PMCID: PMC11093588
- DOI: 10.1126/sciimmunol.adg1094
TIM-3+ CD8 T cells with a terminally exhausted phenotype retain functional capacity in hematological malignancies
Abstract
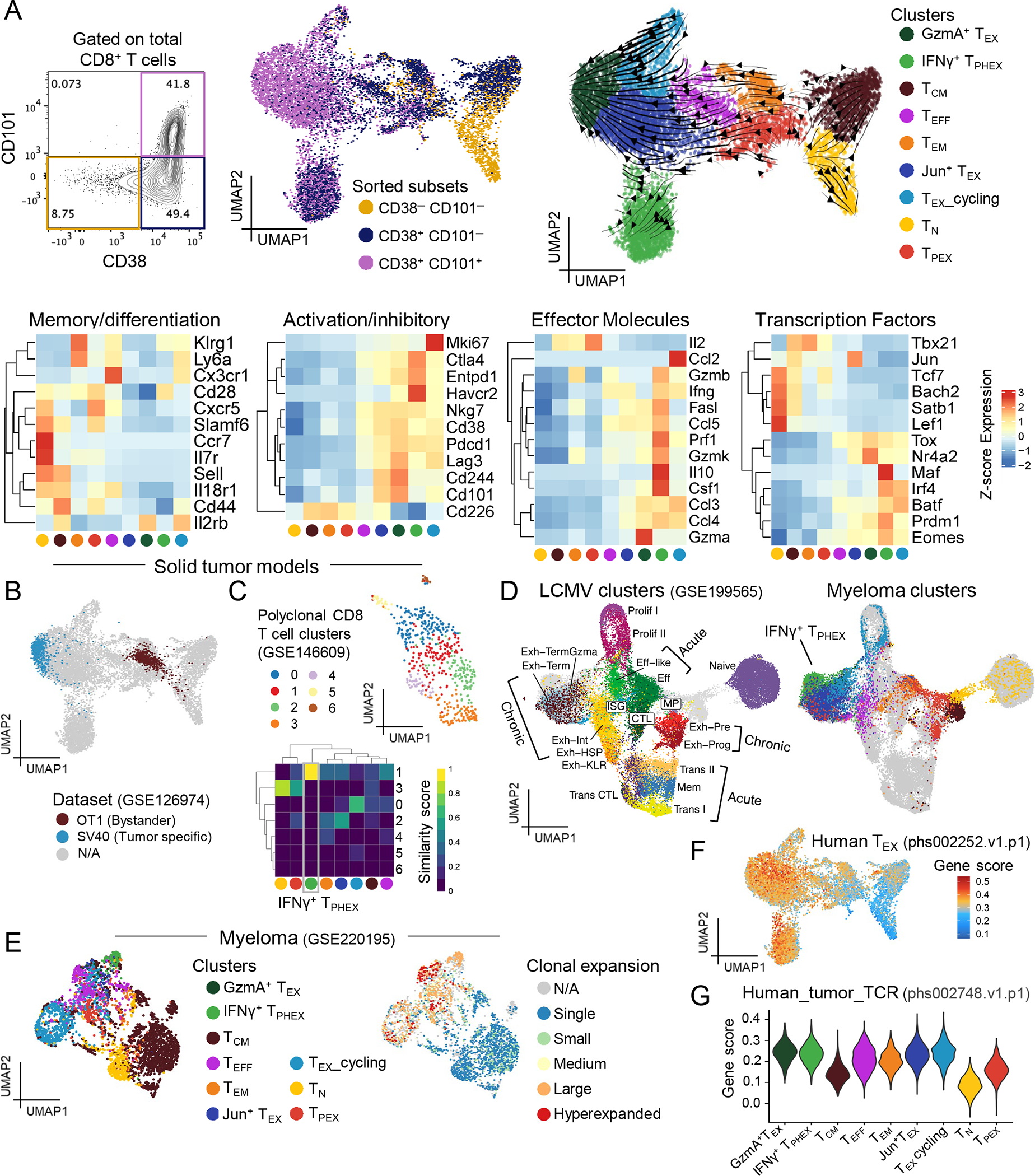
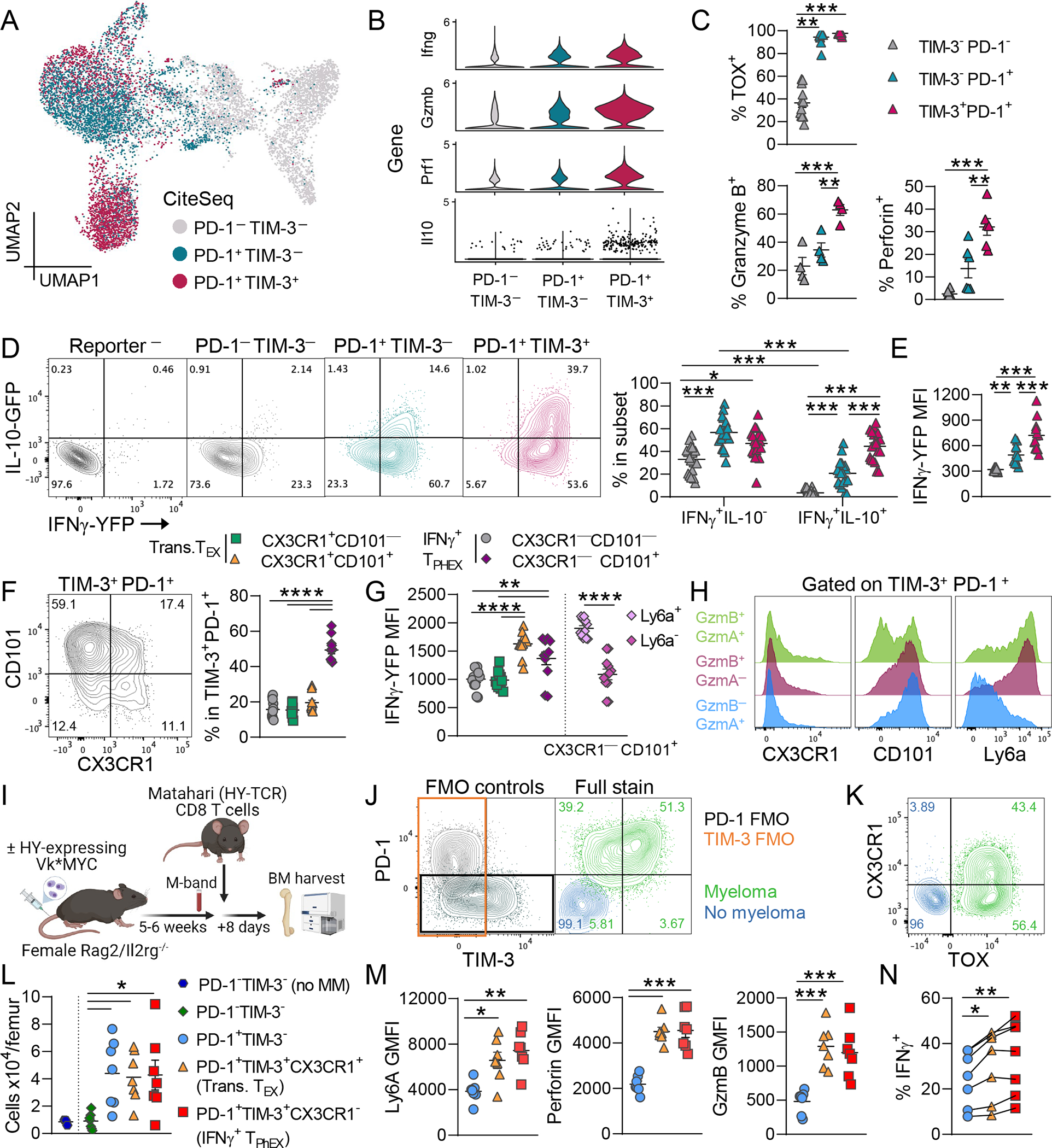
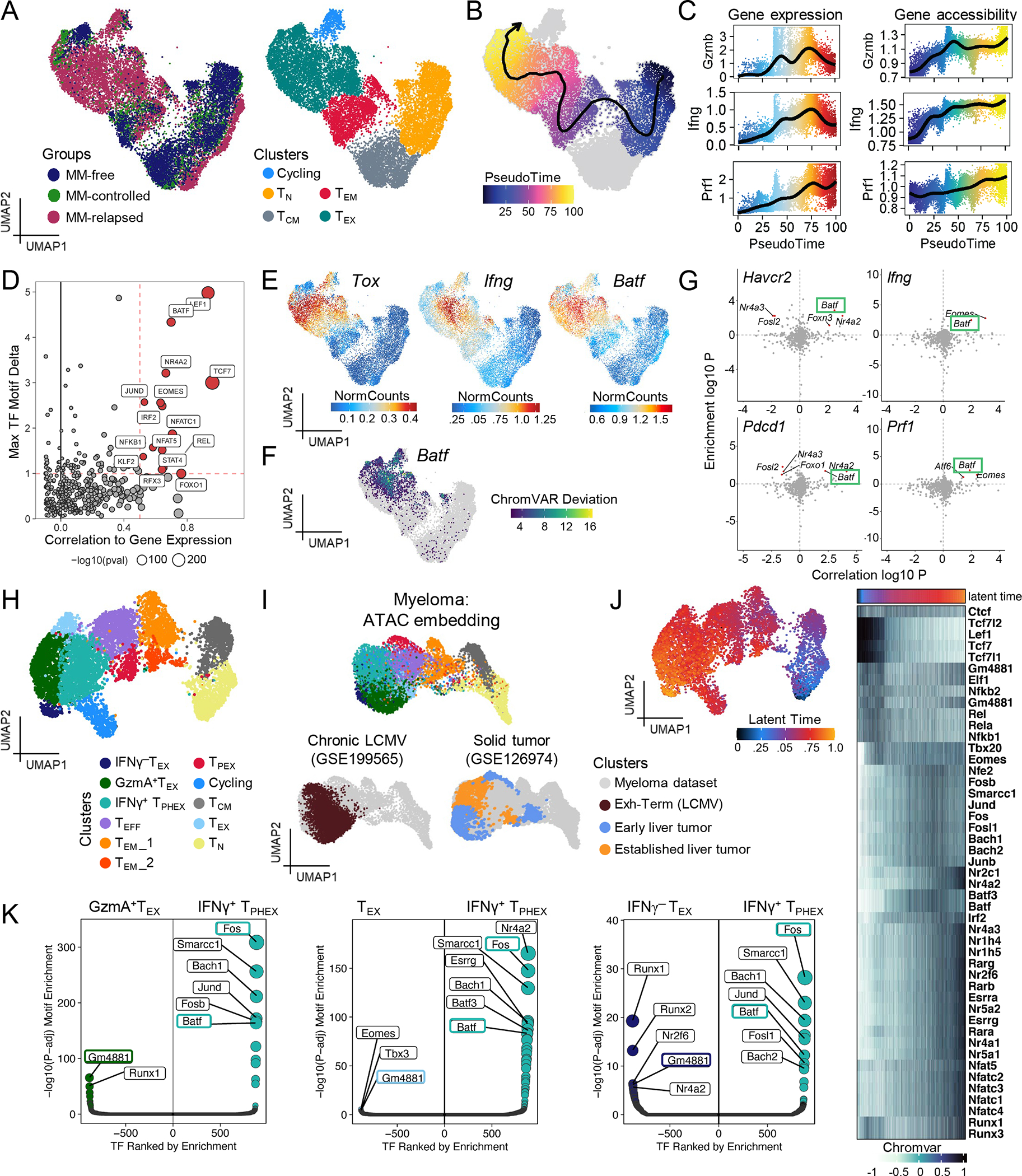
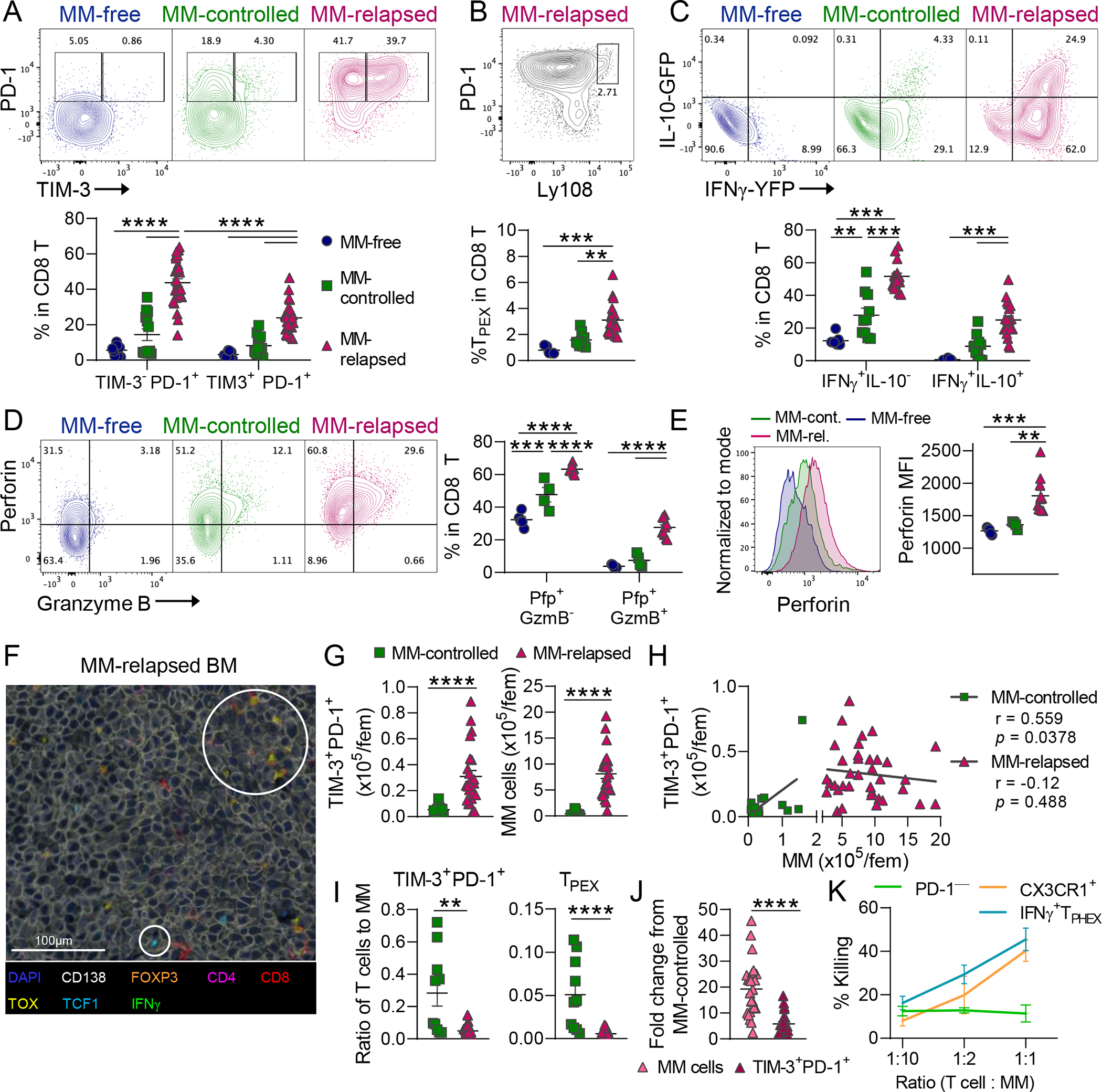
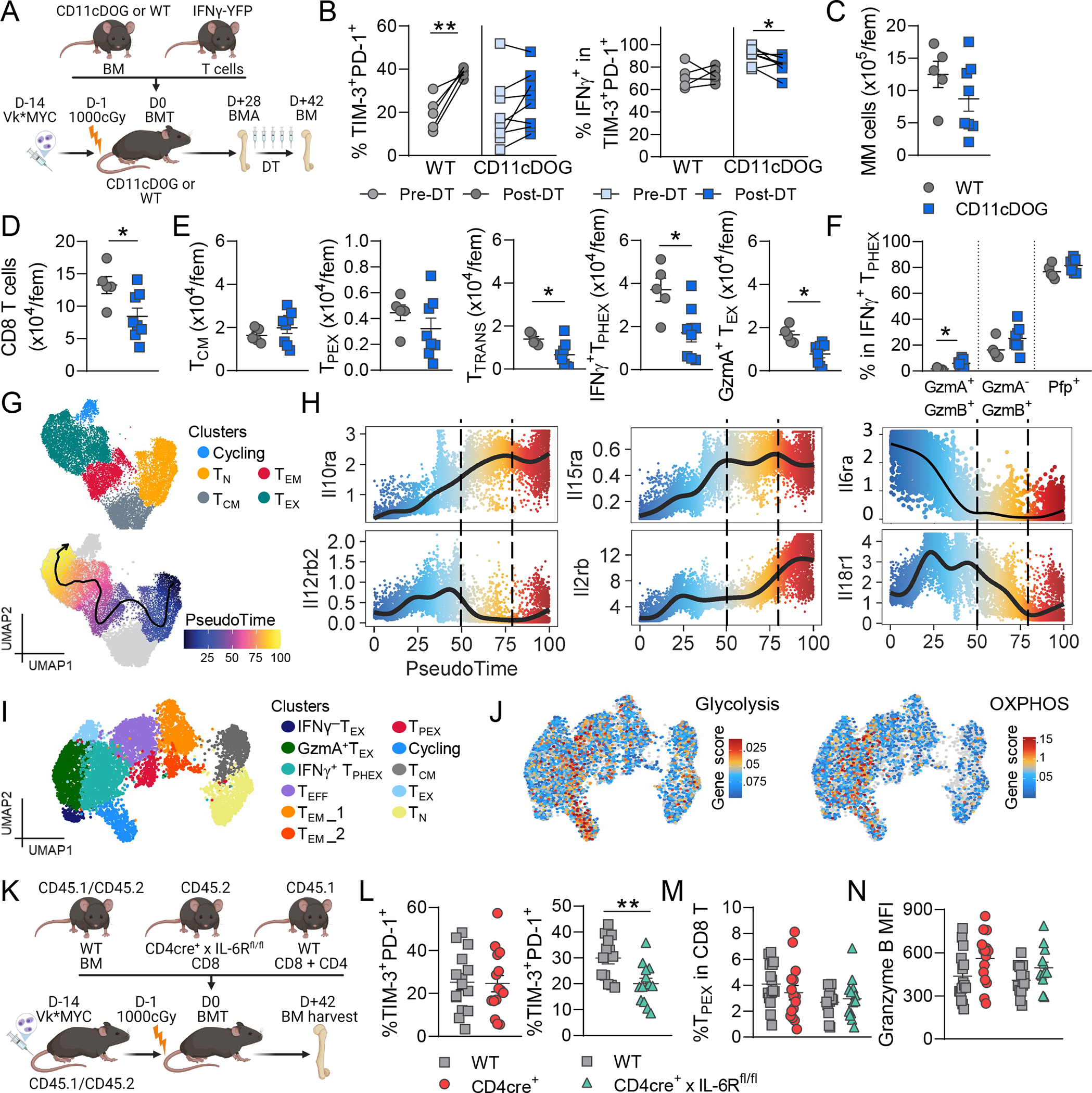
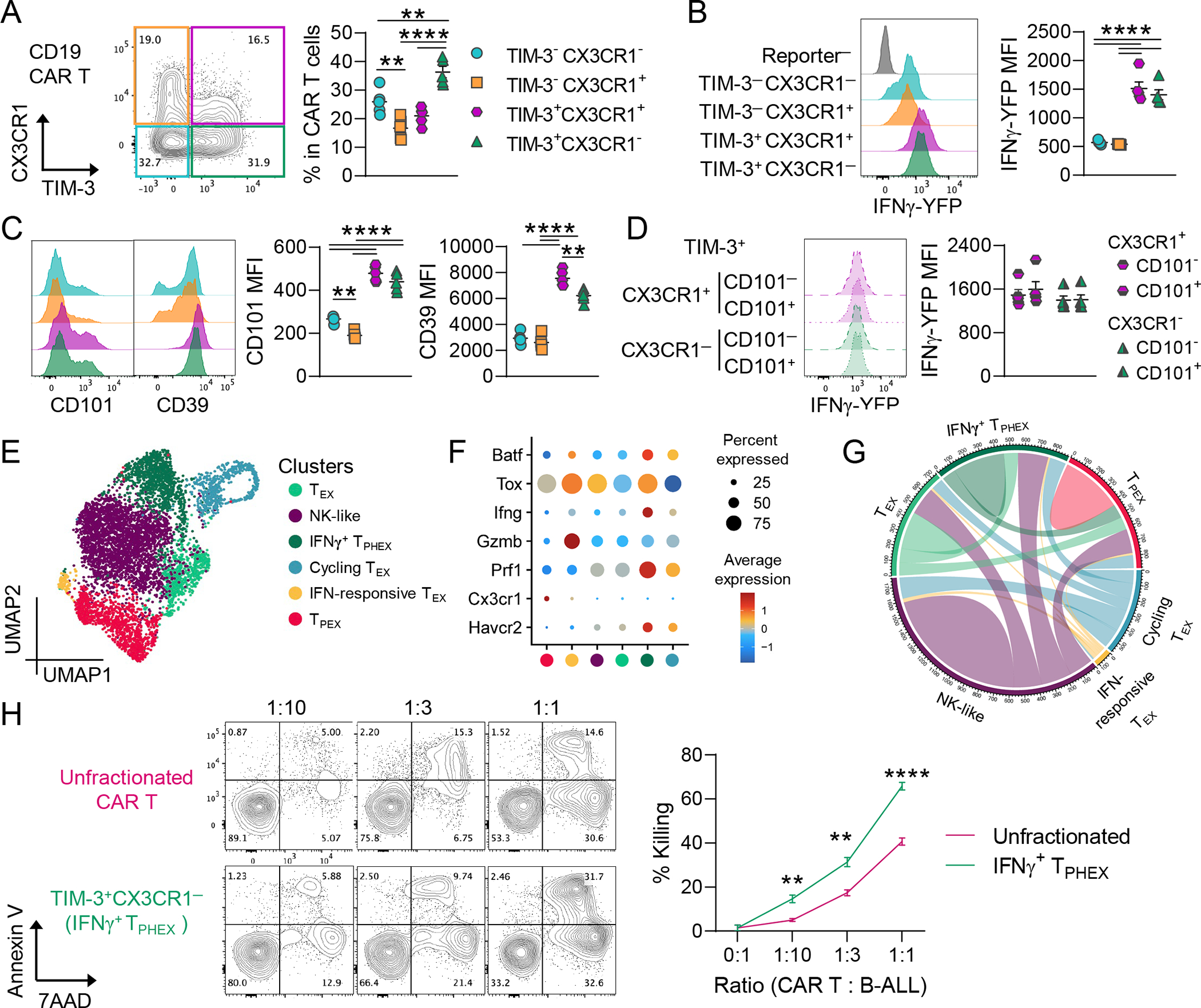
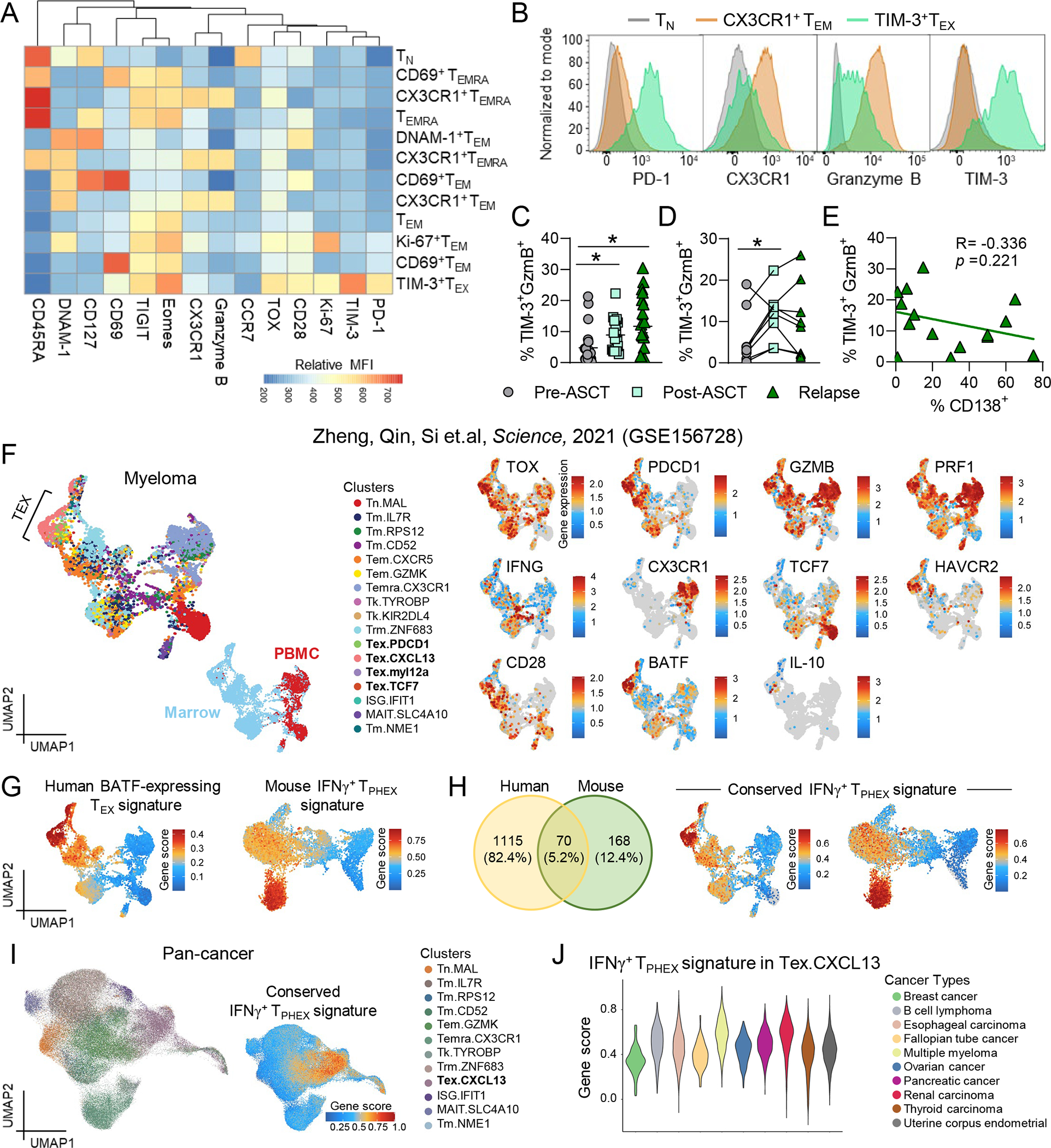
Chronic antigen stimulation is thought to generate dysfunctional CD8 T cells. Here, we identify a CD8 T cell subset in the bone marrow tumor microenvironment that, despite an apparent terminally exhausted phenotype (TPHEX), expressed granzymes, perforin, and IFN-γ. Concurrent gene expression and DNA accessibility revealed that genes encoding these functional proteins correlated with BATF expression and motif accessibility. IFN-γ+ TPHEX effectively killed myeloma with comparable efficacy to transitory effectors, and disease progression correlated with numerical deficits in IFN-γ+ TPHEX. We also observed IFN-γ+ TPHEX within CD19-targeted chimeric antigen receptor T cells, which killed CD19+ leukemia cells. An IFN-γ+ TPHEX gene signature was recapitulated in TEX cells from human cancers, including myeloma and lymphoma. Here, we characterize a TEX subset in hematological malignancies that paradoxically retains function and is distinct from dysfunctional TEX found in chronic viral infections. Thus, IFN-γ+ TPHEX represent a potential target for immunotherapy of blood cancers.
Conflict of interest statement
Competing Interests:
SRR is a founder and shareholder of Lyell Immunopharma and Juno Therapeutics, a Bristol Myers Squibb company. SRR serves as an advisor to Lyell Immunopharma and Adaptive Biotechnologies, has intellectual property licensed to Lyell Immunopharma and Juno/BMS, and receives research support from Lyell Immunopharma and Bristol Myers Squibb. DJG has received research funding, has served as an advisor and has received royalties from Juno Therapeutics, a Bristol-Myers Squibb company; has served as an advisor and received research funding from Seattle Genetics; has served as an advisor for GlaxoSmithKline, Celgene, Janssen Biotech, Ensoma and Legend Biotech; and has received research funding from SpringWorks Therapeutics, Sanofi, and Cellectar Biosciences. GRH has consulted for Generon Corporation, NapaJen Pharma, iTeos Therapeutics, Neoleukin Therapeutics, CSL Behring, Cynata Therapeutics and has received research funding from Compass Therapeutics, Syndax Pharmaceuticals, Applied Molecular Transport, Serplus Technology, Heat Biologics, Laevoroc Oncology and iTeos Therapeutics. All other authors declare that they have no competing interests.
Figures
Similar articles
-
CXCR5 and TIM-3 expressions define distinct exhausted T cell subsets in experimental cutaneous infection with Leishmania mexicana.Front Immunol. 2023 Aug 25;14:1231836. doi: 10.3389/fimmu.2023.1231836. eCollection 2023. Front Immunol. 2023. PMID: 37691941 Free PMC article.
-
Impact of CD4 T cells on intratumoral CD8 T-cell exhaustion and responsiveness to PD-1 blockade therapy in mouse brain tumors.J Immunother Cancer. 2022 Dec;10(12):e005293. doi: 10.1136/jitc-2022-005293. J Immunother Cancer. 2022. PMID: 36543376 Free PMC article.
-
Increased exhausted CD8+ T cells with programmed death-1, T-cell immunoglobulin and mucin-domain-containing-3 phenotype in patients with multiple myeloma.Asia Pac J Clin Oncol. 2018 Oct;14(5):e266-e274. doi: 10.1111/ajco.13033. Epub 2018 Jun 25. Asia Pac J Clin Oncol. 2018. PMID: 29943497
-
The tumor microenvironment shapes the molecular characteristics of exhausted CD8+ T cells.Cancer Lett. 2021 May 28;506:55-66. doi: 10.1016/j.canlet.2021.02.013. Epub 2021 Mar 2. Cancer Lett. 2021. PMID: 33662493 Review.
-
Application of Chimeric Antigen Receptor T Cells in the Treatment of Hematological Malignancies.Biomed Res Int. 2020 Oct 1;2020:4241864. doi: 10.1155/2020/4241864. eCollection 2020. Biomed Res Int. 2020. PMID: 33062678 Free PMC article. Review.
Cited by
-
The Functional and Prognostic Impact of TIGIT Expression on Bone Marrow NK Cells in Core Binding Factor-Acute Myeloid Leukemia Patients at Diagnosis.Biomedicines. 2024 Sep 27;12(10):2207. doi: 10.3390/biomedicines12102207. Biomedicines. 2024. PMID: 39457520 Free PMC article.
-
Bacterial vaginosis-driven changes in vaginal T cell phenotypes and their implications for HIV susceptibility.bioRxiv [Preprint]. 2024 Jul 5:2024.07.03.601916. doi: 10.1101/2024.07.03.601916. bioRxiv. 2024. PMID: 39005354 Free PMC article. Preprint.
-
Hallmarks of tumor-experienced T cells are absent in multiple myeloma patients from diagnosis through maintenance therapy.bioRxiv [Preprint]. 2024 Jun 4:2024.06.03.597178. doi: 10.1101/2024.06.03.597178. bioRxiv. 2024. PMID: 38895348 Free PMC article. Preprint.
-
TOX2 nuclear-cytosol translocation is linked to leukemogenesis of acute T-cell leukemia by repressing TIM3 transcription.Cell Death Differ. 2024 Nov;31(11):1506-1518. doi: 10.1038/s41418-024-01352-z. Epub 2024 Jul 30. Cell Death Differ. 2024. PMID: 39080376 Free PMC article.
-
Cold and hot tumors: from molecular mechanisms to targeted therapy.Signal Transduct Target Ther. 2024 Oct 18;9(1):274. doi: 10.1038/s41392-024-01979-x. Signal Transduct Target Ther. 2024. PMID: 39420203 Free PMC article. Review.
References
-
- Scott AC, Dundar F, Zumbo P, Chandran SS, Klebanoff CA, Shakiba M, Trivedi P, Menocal L, Appleby H, Camara S, Zamarin D, Walther T, Snyder A, Femia MR, Comen EA, Wen HY, Hellmann MD, Anandasabapathy N, Liu Y, Altorki NK, Lauer P, Levy O, Glickman MS, Kaye J, Betel D, Philip M, Schietinger A. TOX is a critical regulator of tumour-specific T cell differentiation. Nature. 2019;571(7764):270–274. - PMC - PubMed
-
- Khan O, Giles JR, McDonald S, Manne S, Ngiow SF, Patel KP, Werner MT, Huang AC, Alexander KA, Wu JE, Attanasio J, Yan P, George SM, Bengsch B, Staupe RP, Donahue G, Xu W, Amaravadi RK, Xu X, Karakousis GC, Mitchell TC, Schuchter LM, Kaye J, Berger SL, Wherry EJ. TOX transcriptionally and epigenetically programs CD8(+) T cell exhaustion. Nature. 2019;571(7764):211–218. - PMC - PubMed
-
- Seo H, Chen J, González-Avalos E, Samaniego-Castruita D, Das A, Wang YH, López-Moyado IF, Georges RO, Zhang W, Onodera A, Wu CJ, Lu LF, Hogan PG, Bhandoola A, Rao A. TOX and TOX2 transcription factors cooperate with NR4A transcription factors to impose CD8(+) T cell exhaustion. Proceedings of the National Academy of Sciences of the United States of America. 2019;116(25):12410–12415. - PMC - PubMed
-
- Yao C, Sun H-W, Lacey NE, Ji Y, Moseman EA, Shih H-Y, Heuston EF, Kirby M, Anderson S, Cheng J, Khan O, Handon R, Reilley J, Fioravanti J, Hu J, Gossa S, Wherry EJ, Gattinoni L, McGavern DB, O’Shea JJ, Schwartzberg PL, Wu T. Single-cell RNA-seq reveals TOX as a key regulator of CD8+ T cell persistence in chronic infection. Nature immunology. 2019;20(7):890–901. - PMC - PubMed
-
- Beltra JC, Manne S, Abdel-Hakeem MS, Kurachi M, Giles JR, Chen Z, Casella V, Ngiow SF, Khan O, Huang YJ, Yan P, Nzingha K, Xu W, Amaravadi RK, Xu X, Karakousis GC, Mitchell TC, Schuchter LM, Huang AC, Wherry EJ. Developmental Relationships of Four Exhausted CD8(+) T Cell Subsets Reveals Underlying Transcriptional and Epigenetic Landscape Control Mechanisms. Immunity. 2020;52(5):825–841.e828. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
