

Building Haplotype-Resolved 3D Genome Maps of Chicken Skeletal Muscle
- PMID: 38582509
- PMCID: PMC11200017
- DOI: 10.1002/advs.202305706
Building Haplotype-Resolved 3D Genome Maps of Chicken Skeletal Muscle
Abstract
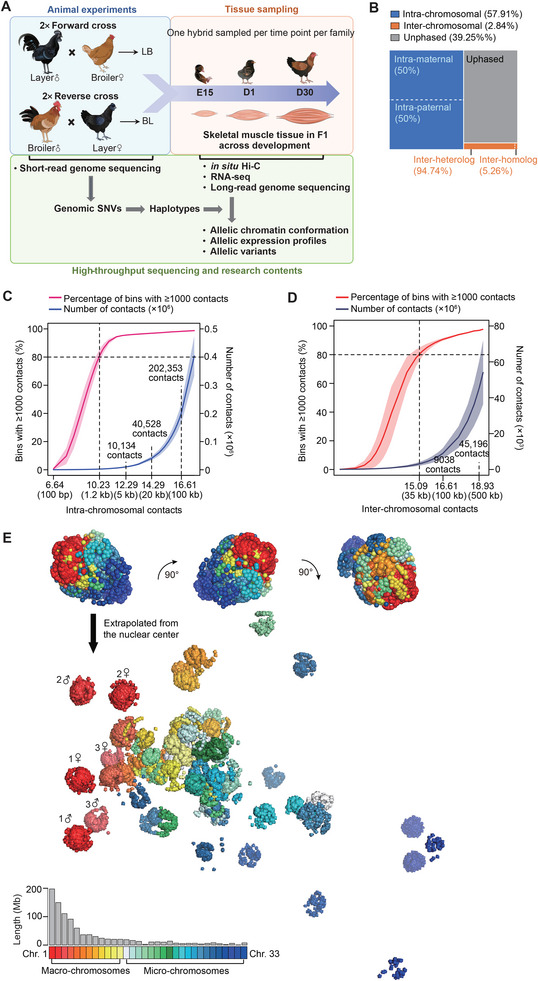
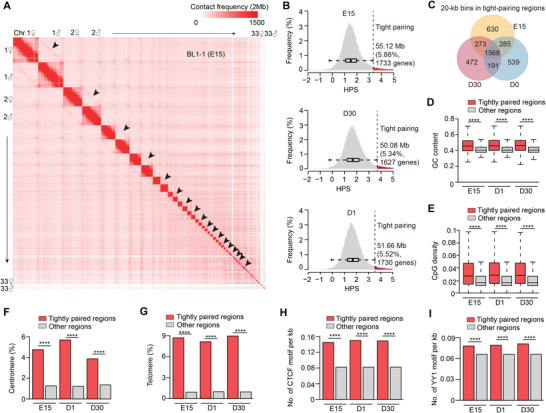
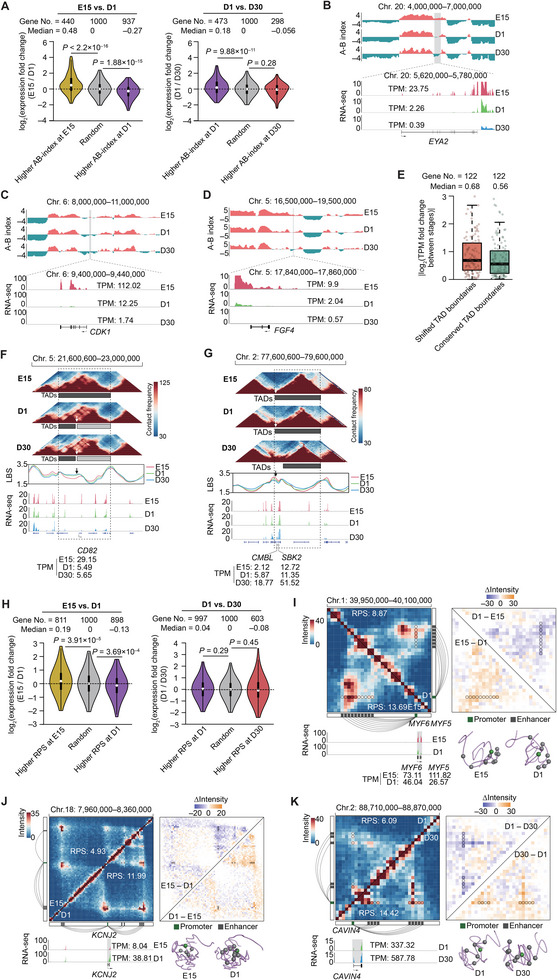
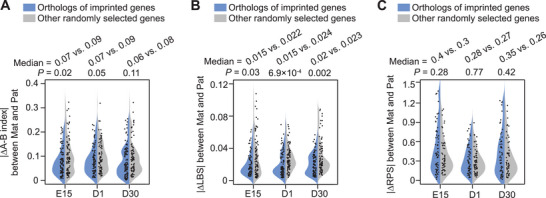
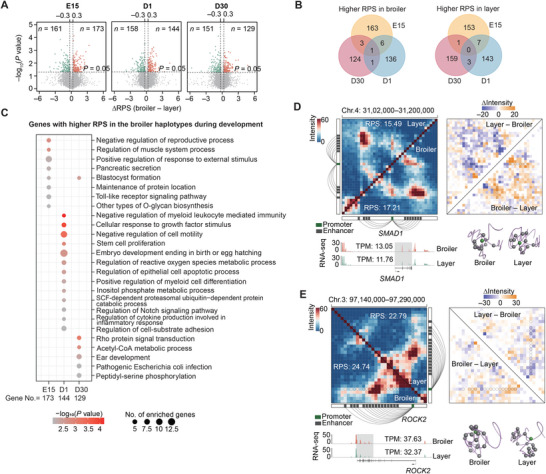
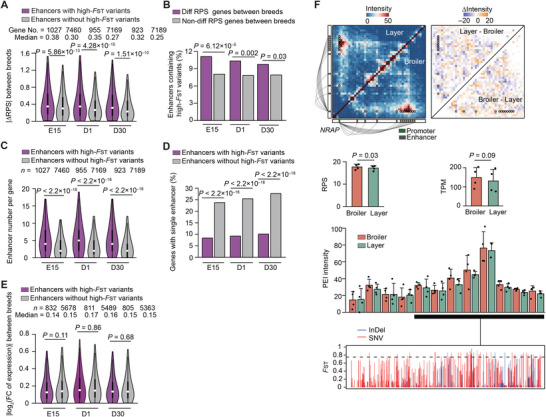
Haplotype-resolved 3D chromatin architecture related to allelic differences in avian skeletal muscle development has not been addressed so far, although chicken husbandry for meat consumption has been prevalent feature of cultures on every continent for more than thousands of years. Here, high-resolution Hi-C diploid maps (1.2-kb maximum resolution) are generated for skeletal muscle tissues in chicken across three developmental stages (embryonic day 15 to day 30 post-hatching). The sequence features governing spatial arrangement of chromosomes and characterize homolog pairing in the nucleus, are identified. Multi-scale characterization of chromatin reorganization between stages from myogenesis in the fetus to myofiber hypertrophy after hatching show concordant changes in transcriptional regulation by relevant signaling pathways. Further interrogation of parent-of-origin-specific chromatin conformation supported that genomic imprinting is absent in birds. This study also reveals promoter-enhancer interaction (PEI) differences between broiler and layer haplotypes in skeletal muscle development-related genes are related to genetic variation between breeds, however, only a minority of breed-specific variations likely contribute to phenotypic divergence in skeletal muscle potentially via allelic PEI rewiring. Beyond defining the haplotype-specific 3D chromatin architecture in chicken, this study provides a rich resource for investigating allelic regulatory divergence among chicken breeds.
Keywords: Hi‐C; chicken; homolog; sequence variations; skeletal muscle development.
© 2024 The Authors. Advanced Science published by Wiley‐VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Haplotype-resolved 3D chromatin architecture of the hybrid pig.Genome Res. 2024 Mar 20;34(2):310-325. doi: 10.1101/gr.278101.123. Genome Res. 2024. PMID: 38479837 Free PMC article.
-
Potential role of miR-155-5p in fat deposition and skeletal muscle development of chicken.Biosci Rep. 2020 Jun 26;40(6):BSR20193796. doi: 10.1042/BSR20193796. Biosci Rep. 2020. PMID: 32441300 Free PMC article.
-
RNA sequencing for global gene expression associated with muscle growth in a single male modern broiler line compared to a foundational Barred Plymouth Rock chicken line.BMC Genomics. 2017 Jan 13;18(1):82. doi: 10.1186/s12864-016-3471-y. BMC Genomics. 2017. PMID: 28086790 Free PMC article.
-
Understanding three-dimensional chromatin organization in diploid genomes.Comput Struct Biotechnol J. 2021 Jun 15;19:3589-3598. doi: 10.1016/j.csbj.2021.06.018. eCollection 2021. Comput Struct Biotechnol J. 2021. PMID: 34257838 Free PMC article. Review.
-
A (3D-Nuclear) Space Odyssey: Making Sense of Hi-C Maps.Genes (Basel). 2019 May 29;10(6):415. doi: 10.3390/genes10060415. Genes (Basel). 2019. PMID: 31146487 Free PMC article. Review.
References
-
- a) Asp P., Blum R., Vethantham V., Parisi F., Micsinai M., Cheng J., Bowman C., Kluger Y., Dynlacht B. D., Natl. Acad. Sci. USA. 2011, 108, E149; - PMC - PubMed
- b) Buckingham M., Curr. Opin. Genet. Dev. 2001, 11, 440; - PubMed
- c) Bentzinger C. F., Wang Y. X., Rudnicki M. A., Cold Spring Harbor Perspect. Biol. 2012, 4, a008342; - PMC - PubMed
- d) Wang R., Chen F., Chen Q., Wan X., Shi M., Chen A. K., Ma Z., Li G., Wang M., Ying Y., Liu Q., Li H., Zhang X., Ma J., Zhong J., Chen M., Zhang M. Q., Zhang Y., Chen Y., Zhu D., Nat. Commun. 2022, 13, 205. - PMC - PubMed
-
- a) Yu M., Wang H., Xu Y., Yu D., Li D., Liu X., Du W., Cell Biol Int 2015, 39, 910; - PubMed
- b) Chen M. M., Zhao Y. P., Zhao Y., Deng S. L., Yu K., Front Cell Dev Biol 2021, 9, 785712; - PMC - PubMed
- c) Shirakawa T., Toyono T., Inoue A., Matsubara T., Kawamoto T., Kokabu S., Cells 2022, 11, 1493. - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources