

Co-imaging of RelA and c-Rel reveals features of NF-κB signaling for ligand discrimination
- PMID: 38483906
- PMCID: PMC11015162
- DOI: 10.1016/j.celrep.2024.113940
Co-imaging of RelA and c-Rel reveals features of NF-κB signaling for ligand discrimination
Abstract
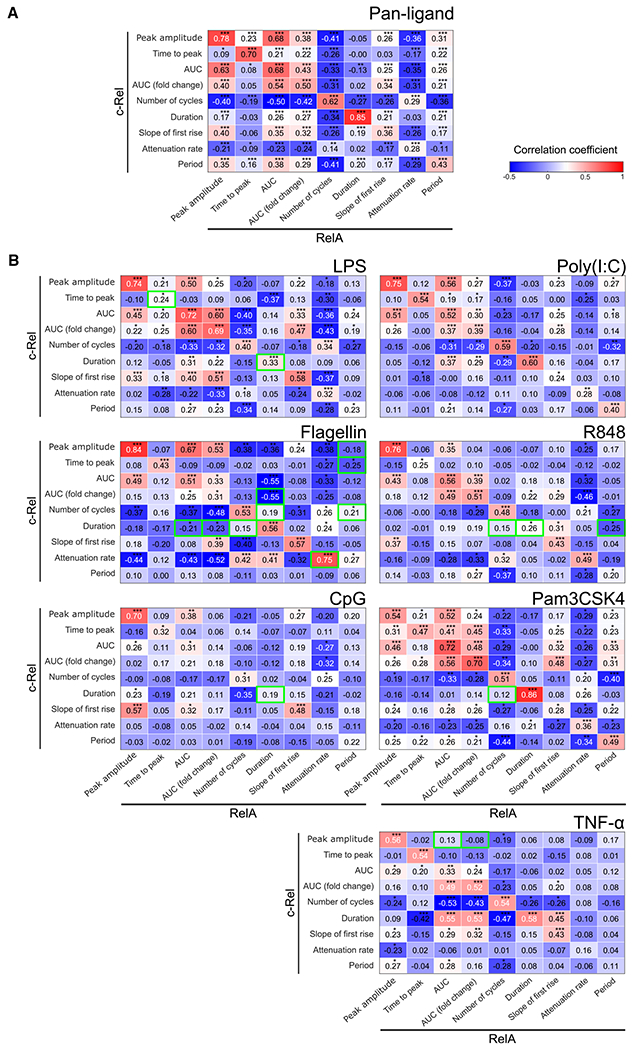
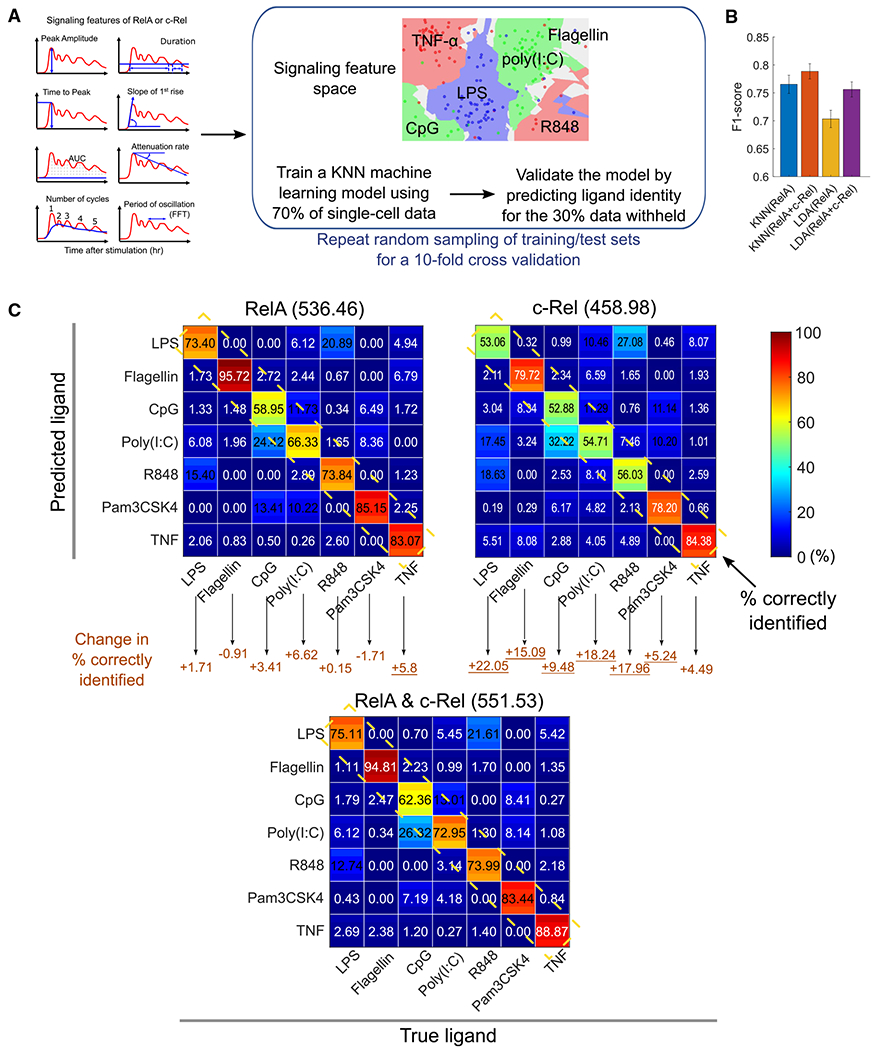
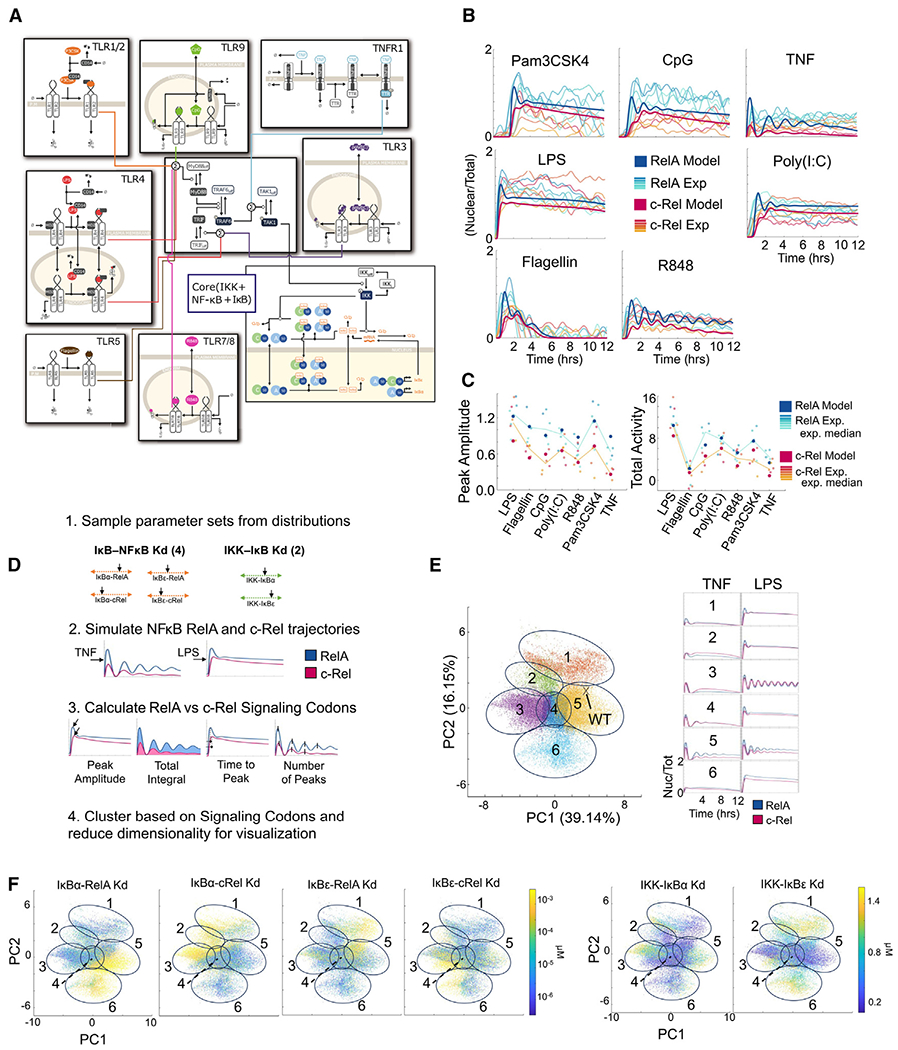
Individual cell sensing of external cues has evolved through the temporal patterns in signaling. Since nuclear factor κB (NF-κB) signaling dynamics have been examined using a single subunit, RelA, it remains unclear whether more information might be transmitted via other subunits. Using NF-κB double-knockin reporter mice, we monitored both canonical NF-κB subunits, RelA and c-Rel, simultaneously in single macrophages by quantitative live-cell imaging. We show that signaling features of RelA and c-Rel convey more information about the stimuli than those of either subunit alone. Machine learning is used to predict the ligand identity accurately based on RelA and c-Rel signaling features without considering the co-activated factors. Ligand discrimination is achieved through selective non-redundancy of RelA and c-Rel signaling dynamics, as well as their temporal coordination. These results suggest a potential role of c-Rel in fine-tuning immune responses and highlight the need for approaches that will elucidate the mechanisms regulating NF-κB subunit specificity.
Keywords: CP: Molecular biology; NF-κB; RelA; c-Rel; endogenous knockin; fluorescent fusion reporter mice; inflammatory signaling; live microscopy; macrophages; mathematical modeling.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures





Similar articles
-
Integrative analysis suggests cell type-specific decoding of NF-κB dynamics.Sci Signal. 2020 Feb 25;13(620):eaax7195. doi: 10.1126/scisignal.aax7195. Sci Signal. 2020. PMID: 32098801 Free PMC article.
-
Double knockin mice show NF-κB trajectories in immune signaling and aging.Cell Rep. 2022 Nov 22;41(8):111682. doi: 10.1016/j.celrep.2022.111682. Cell Rep. 2022. PMID: 36417863 Free PMC article.
-
Differential requirements for the canonical NF-κB transcription factors c-REL and RELA during the generation and activation of mature B cells.Immunol Cell Biol. 2017 Mar;95(3):261-271. doi: 10.1038/icb.2016.95. Epub 2016 Sep 21. Immunol Cell Biol. 2017. PMID: 27649781 Free PMC article.
-
Rel/NF-kappaB transcription factors: key mediators of B-cell activation.Immunol Rev. 2000 Aug;176:134-40. doi: 10.1034/j.1600-065x.2000.00615.x. Immunol Rev. 2000. PMID: 11043773 Review.
-
Aberrant rel/nfkb genes and activity in human cancer.Oncogene. 1999 Nov 22;18(49):6938-47. doi: 10.1038/sj.onc.1203221. Oncogene. 1999. PMID: 10602468 Review.
Cited by
-
Exploration of the Shared Gene Signatures and Molecular Mechanisms between Chronic Bronchitis and Antineutrophil Cytoplasmic Antibody-associated Glomerulonephritis: Evidence from Transcriptome Data.Curr Pharm Des. 2024;30(25):1966-1984. doi: 10.2174/0113816128297623240521070426. Curr Pharm Des. 2024. PMID: 38847168
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
