

Tools for live-cell imaging of cytoskeletal and nuclear behavior in the unconventional yeast, Aureobasidium pullulans
- PMID: 38446617
- PMCID: PMC11064661
- DOI: 10.1091/mbc.E23-10-0388
Tools for live-cell imaging of cytoskeletal and nuclear behavior in the unconventional yeast, Aureobasidium pullulans
Abstract
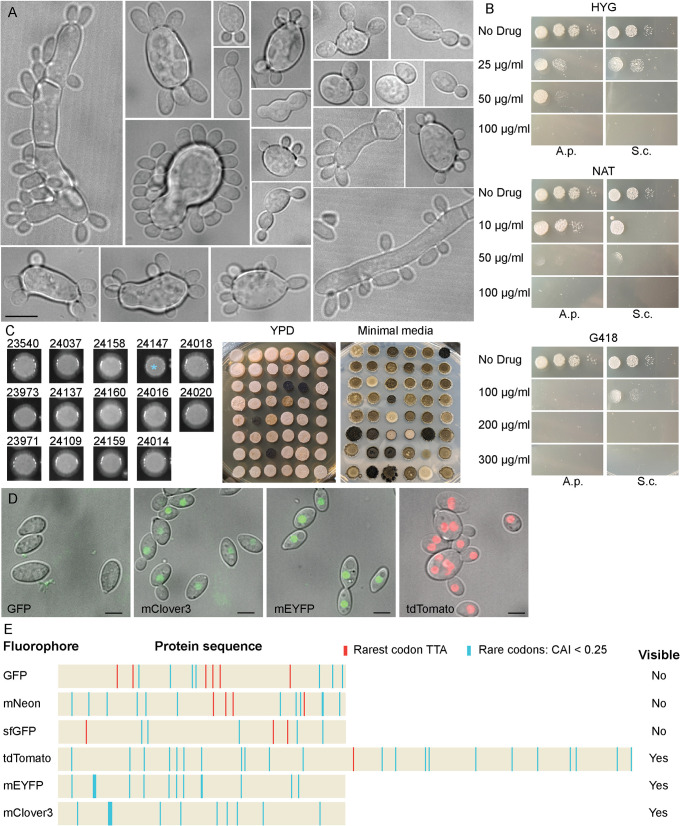
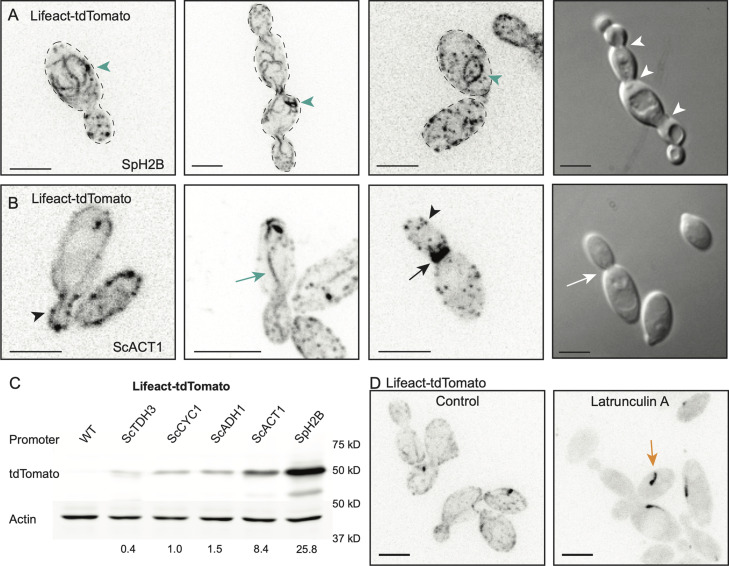
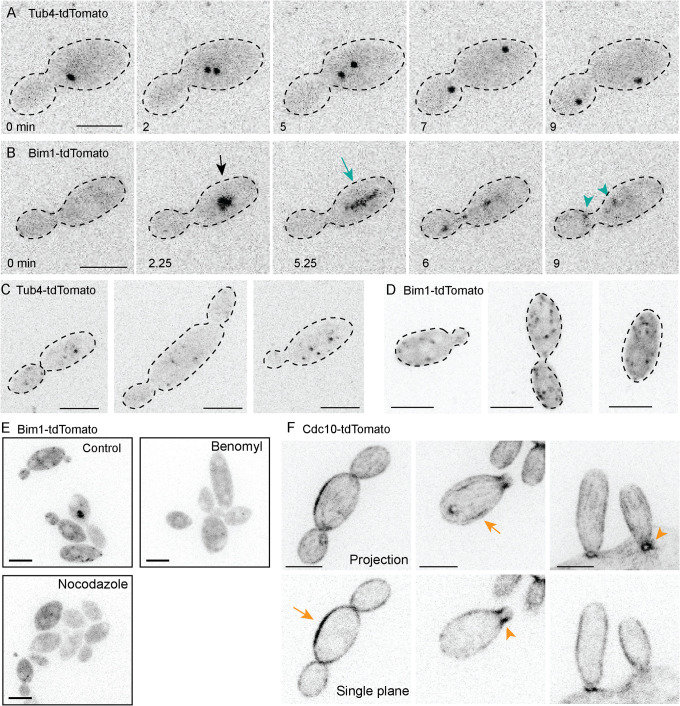
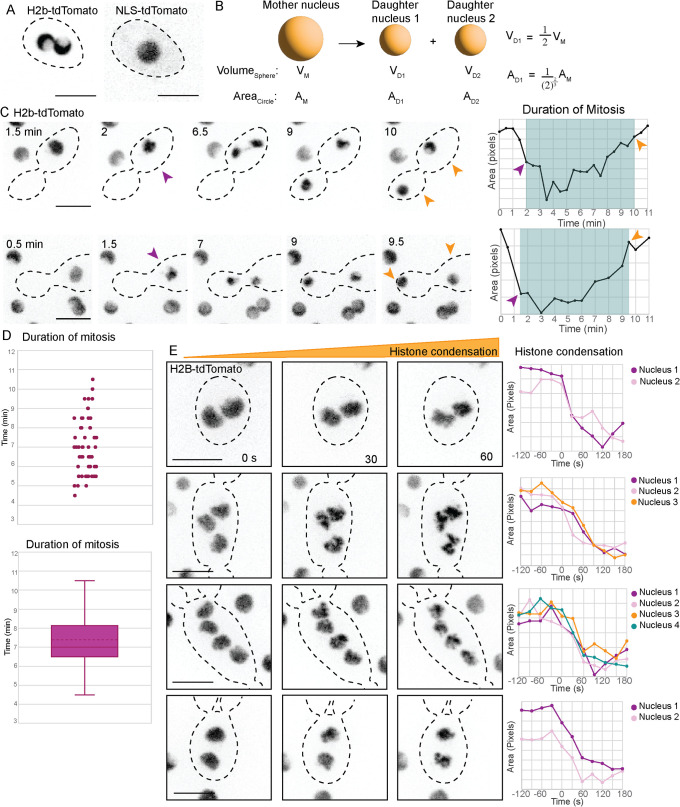
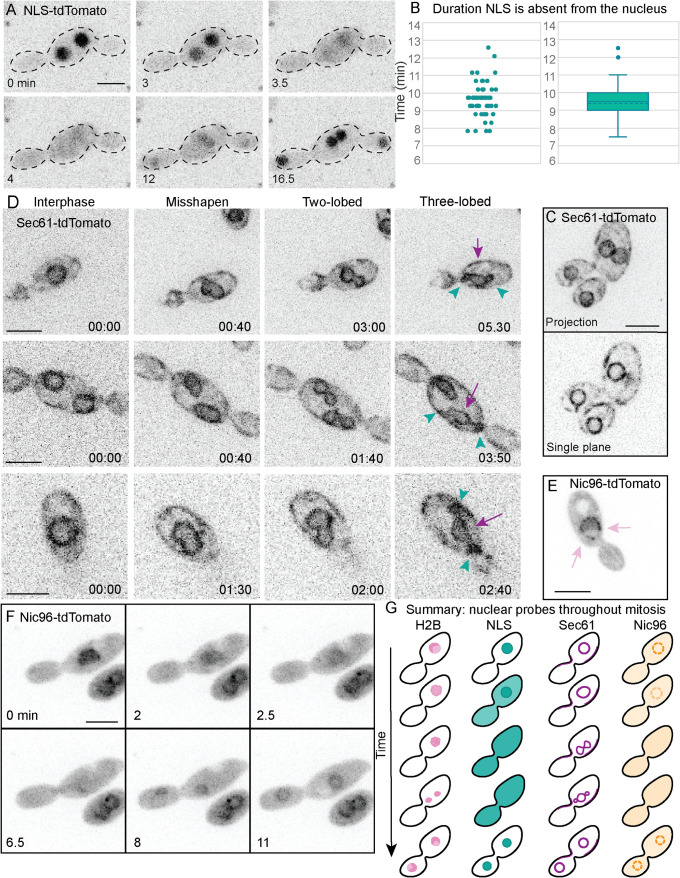
Aureobasidium pullulans is a ubiquitous fungus with a wide variety of morphologies and growth modes including "typical" single-budding yeast, and interestingly, larger multinucleate yeast than can make multiple buds in a single cell cycle. The study of A. pullulans promises to uncover novel cell biology, but currently tools are lacking to achieve this goal. Here, we describe initial components of a cell biology toolkit for A. pullulans, which is used to express and image fluorescent probes for nuclei as well as components of the cytoskeleton. These tools allowed live-cell imaging of the multinucleate and multibudding cycles, revealing highly synchronous mitoses in multinucleate yeast that occur in a semiopen manner with an intact but permeable nuclear envelope. These findings open the door to using this ubiquitous polyextremotolerant fungus as a model for evolutionary cell biology.
Figures
Similar articles
-
Fifty Aureobasidium pullulans genomes reveal a recombining polyextremotolerant generalist.Environ Microbiol. 2019 Oct;21(10):3638-3652. doi: 10.1111/1462-2920.14693. Epub 2019 Jun 18. Environ Microbiol. 2019. PMID: 31112354 Free PMC article.
-
The current status of Aureobasidium pullulans in biotechnology.Folia Microbiol (Praha). 2018 Mar;63(2):129-140. doi: 10.1007/s12223-017-0561-4. Epub 2017 Oct 27. Folia Microbiol (Praha). 2018. PMID: 29079936 Review.
-
Assembly and comparative genome analysis of a Patagonian Aureobasidium pullulans isolate reveals unexpected intraspecific variation.Yeast. 2023 May;40(5-6):197-213. doi: 10.1002/yea.3853. Epub 2023 Apr 28. Yeast. 2023. PMID: 37114349
-
Ferrous chloride and ferrous sulfate improve the fungicidal efficacy of cold atmospheric argon plasma on melanized Aureobasidium pullulans.J Biosci Bioeng. 2019 Jul;128(1):28-32. doi: 10.1016/j.jbiosc.2018.12.008. Epub 2019 Jan 22. J Biosci Bioeng. 2019. PMID: 30679111
-
Bioproducts from Aureobasidium pullulans, a biotechnologically important yeast.Appl Microbiol Biotechnol. 2009 Apr;82(5):793-804. doi: 10.1007/s00253-009-1882-2. Epub 2009 Feb 7. Appl Microbiol Biotechnol. 2009. PMID: 19198830 Review.
Cited by
-
Chemical transformation of the multibudding yeast, Aureobasidium pullulans.J Cell Biol. 2024 Oct 7;223(10):e202402114. doi: 10.1083/jcb.202402114. Epub 2024 Jun 27. J Cell Biol. 2024. PMID: 38935076 Free PMC article.
References
-
- Aoki K, Hayashi H, Furuya K, Sato M, Takagi T, Osumi M, Kimura A, Niki H (2011). Breakage of the nuclear envelope by an extending mitotic nucleus occurs during anaphase in Schizosaccharomyces japonicus. Gene Cells 16, 911–926. - PubMed
MeSH terms
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
