

The antagonistic transcription factors, EspM and EspN, regulate the ESX-1 secretion system in M. marinum
- PMID: 38445877
- PMCID: PMC11005418
- DOI: 10.1128/mbio.03357-23
The antagonistic transcription factors, EspM and EspN, regulate the ESX-1 secretion system in M. marinum
Abstract
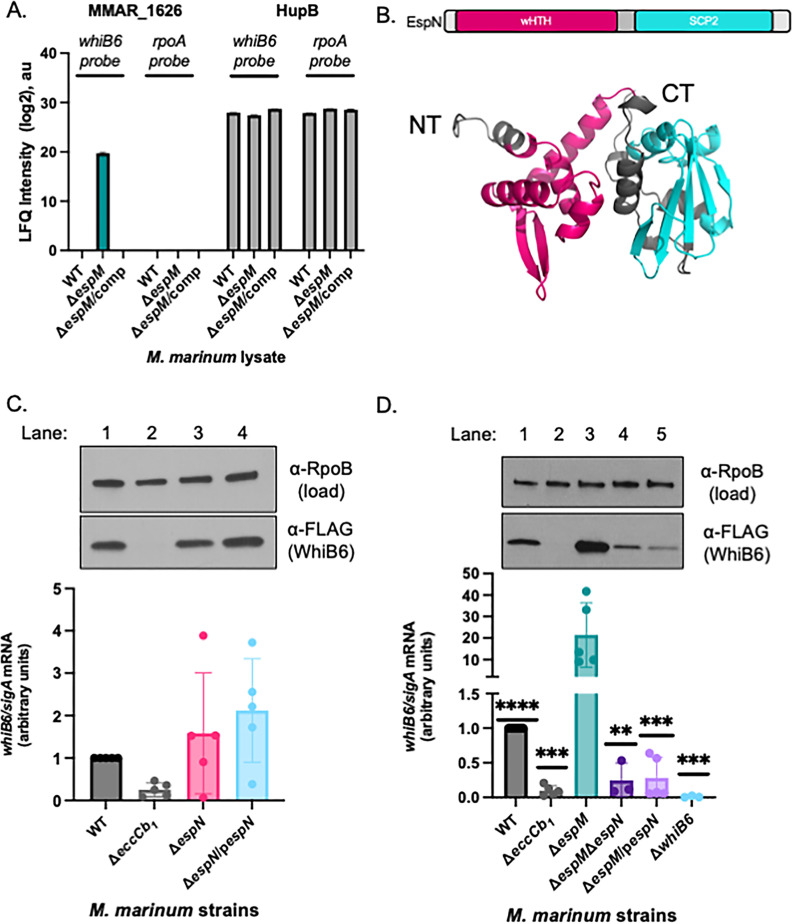
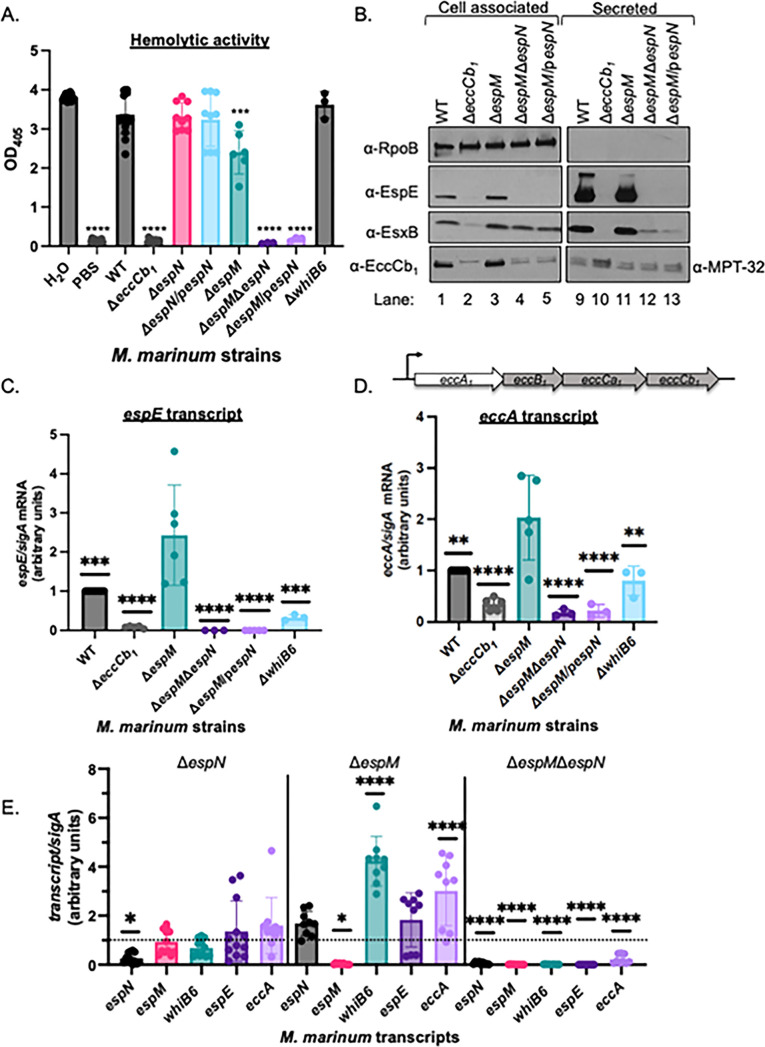
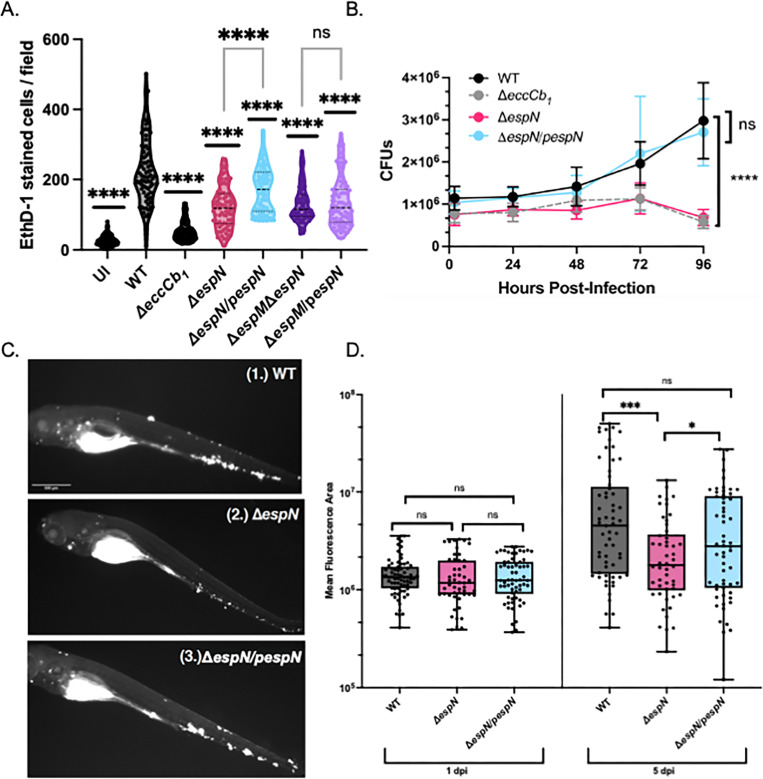
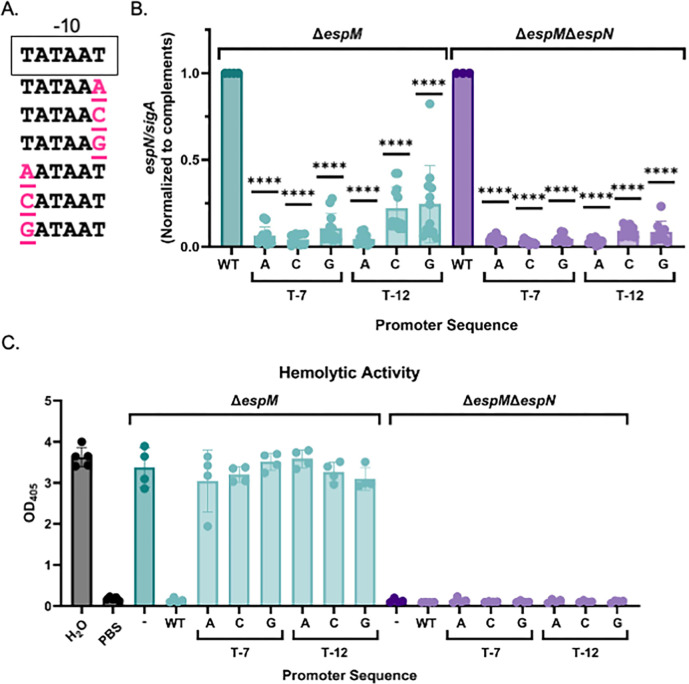
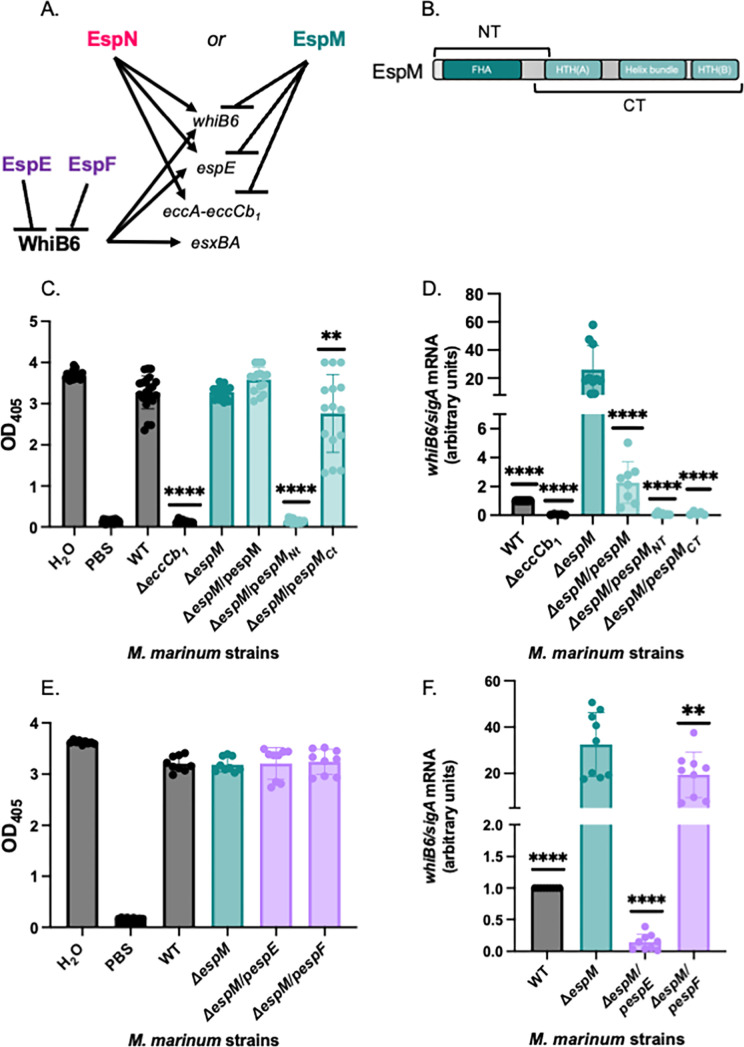
Bacterial pathogens use protein secretion systems to transport virulence factors and regulate gene expression. Among pathogenic mycobacteria, including Mycobacterium tuberculosis and Mycobacterium marinum, the ESAT-6 system 1 (ESX-1) secretion is crucial for host interaction. Secretion of protein substrates by the ESX-1 secretion system disrupts phagosomes, allowing mycobacteria cytoplasmic access during macrophage infections. Deletion or mutation of the ESX-1 system attenuates mycobacterial pathogens. Pathogenic mycobacteria respond to the presence or absence of the ESX-1 system in the cytoplasmic membrane by altering transcription. Under laboratory conditions, the EspM repressor and WhiB6 activator control transcription of specific ESX-1-responsive genes, including the ESX-1 substrate genes. However, deleting the espM or whiB6 gene does not phenocopy the deletion of the ESX-1 substrate genes during macrophage infection by M. marinum. In this study, we identified EspN, a critical transcription factor whose activity is masked by the EspM repressor under laboratory conditions. In the absence of EspM, EspN activates transcription of whiB6 and ESX-1 genes during both laboratory growth and macrophage infection. EspN is also independently required for M. marinum growth within and cytolysis of macrophages, similar to the ESX-1 genes, and for disease burden in a zebrafish larval model of infection. These findings suggest that EspN and EspM coordinate to counterbalance the regulation of the ESX-1 system and support mycobacterial pathogenesis.IMPORTANCEPathogenic mycobacteria, which are responsible for tuberculosis and other long-term diseases, use the ESX-1 system to transport proteins that control the host response to infection and promote bacterial survival. In this study, we identify an undescribed transcription factor that controls the expression of ESX-1 genes and is required for both macrophage and animal infection. However, this transcription factor is not the primary regulator of ESX-1 genes under standard laboratory conditions. These findings identify a critical transcription factor that likely controls expression of a major virulence pathway during infection, but whose effect is not detectable with standard laboratory strains and growth conditions.
Keywords: ESX-1; Mycobacterium; regulation; transcription; type VII secretion.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The EspN transcription factor is an infection-dependent regulator of the ESX-1 system in M. marinum.bioRxiv [Preprint]. 2023 Feb 16:2023.02.15.528779. doi: 10.1101/2023.02.15.528779. bioRxiv. 2023. PMID: 36824794 Free PMC article. Preprint.
-
Functional Analysis of EspM, an ESX-1-Associated Transcription Factor in Mycobacterium marinum.J Bacteriol. 2022 Dec 20;204(12):e0023322. doi: 10.1128/jb.00233-22. Epub 2022 Nov 30. J Bacteriol. 2022. PMID: 36448785 Free PMC article.
-
EspM Is a Conserved Transcription Factor That Regulates Gene Expression in Response to the ESX-1 System.mBio. 2020 Feb 4;11(1):e02807-19. doi: 10.1128/mBio.02807-19. mBio. 2020. PMID: 32019792 Free PMC article.
-
Modeling Tubercular ESX-1 Secretion Using Mycobacterium marinum.Microbiol Mol Biol Rev. 2020 Sep 2;84(4):e00082-19. doi: 10.1128/MMBR.00082-19. Print 2020 Nov 18. Microbiol Mol Biol Rev. 2020. PMID: 32878966 Free PMC article. Review.
-
ESX-3 secretion system in Mycobacterium: An overview.Biochimie. 2024 Jan;216:46-55. doi: 10.1016/j.biochi.2023.10.013. Epub 2023 Oct 23. Biochimie. 2024. PMID: 37879428 Review.
Cited by
-
The regulatory functions of ESX-1 substrates, EspE and EspF, are separable from secretion.J Bacteriol. 2024 Sep 19;206(9):e0027124. doi: 10.1128/jb.00271-24. Epub 2024 Aug 13. J Bacteriol. 2024. PMID: 39136451 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
