

Depletion of the N6-Methyladenosine (m6A) reader protein IGF2BP3 induces ferroptosis in glioma by modulating the expression of GPX4
- PMID: 38429265
- PMCID: PMC10907351
- DOI: 10.1038/s41419-024-06486-z
Depletion of the N6-Methyladenosine (m6A) reader protein IGF2BP3 induces ferroptosis in glioma by modulating the expression of GPX4
Abstract
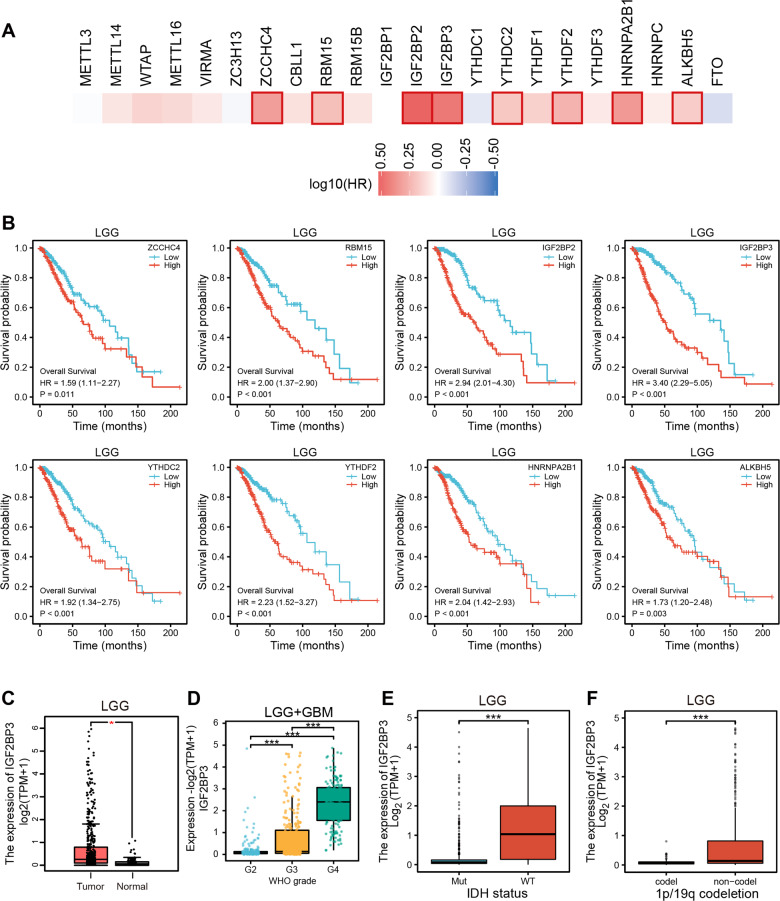
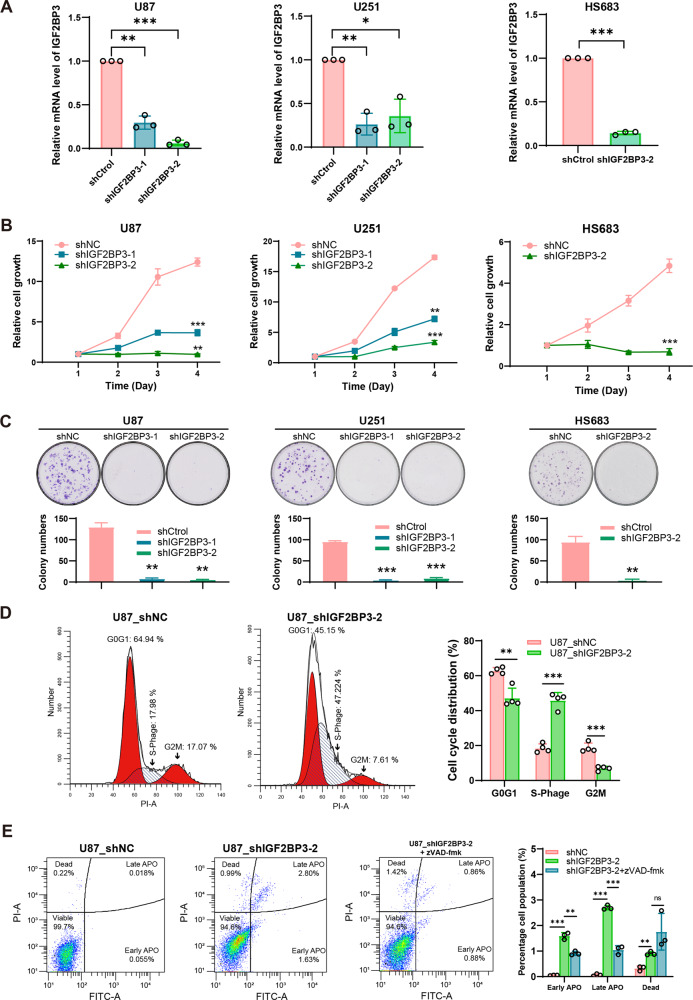
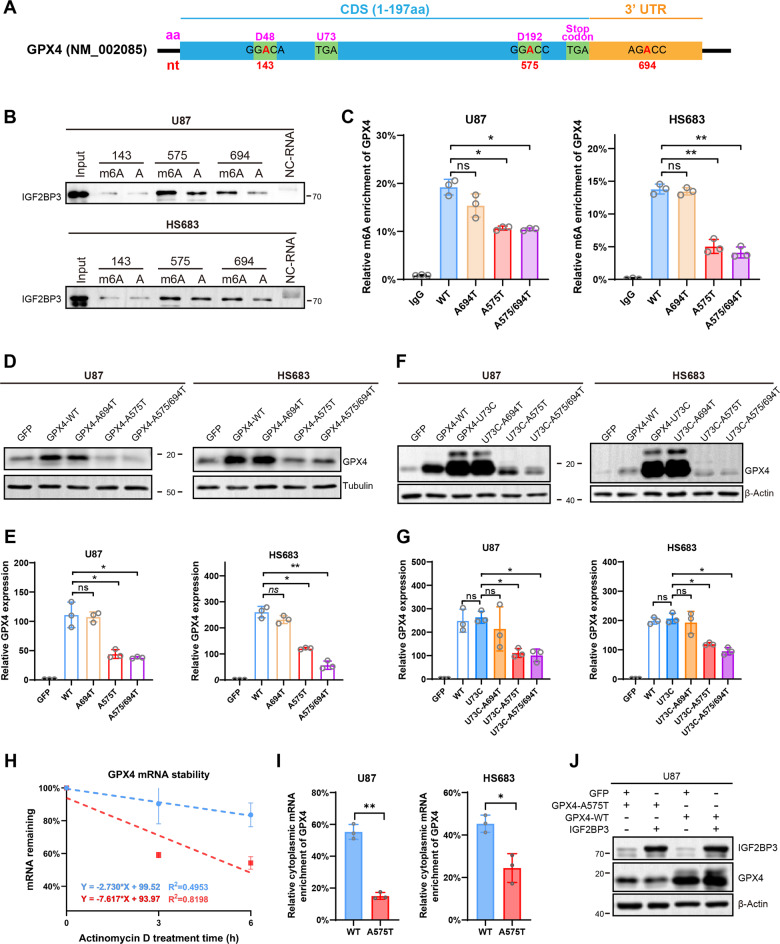
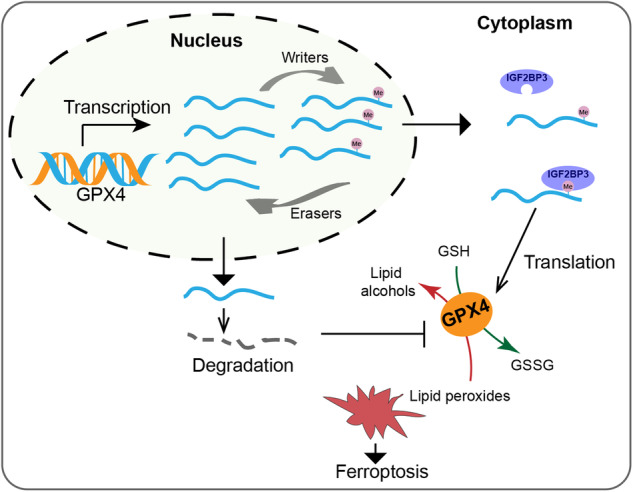
Emerging evidence highlights the multifaceted contributions of m6A modifications to glioma. IGF2BP3, a m6A modification reader protein, plays a crucial role in post-transcriptional gene regulation. Though several studies have identified IGF2BP3 as a poor prognostic marker in glioma, the underlying mechanism remains unclear. In this study, we demonstrated that IGF2BP3 knockdown is detrimental to cell growth and survival in glioma cells. Notably, we discovered that IGF2BP3 regulated ferroptosis by modulating the protein expression level of GPX4 through direct binding to a specific motif on GPX4 mRNA. Strikingly, the m6A modification at this motif was found to be critical for GPX4 mRNA stability and translation. Furthermore, IGF2BP3 knockdown glioma cells were incapable of forming tumors in a mouse xenograft model and were more susceptible to phagocytosis by microglia. Our findings shed light on an unrecognized regulatory function of IGF2BP3 in ferroptosis. The identification of a critical m6A site within the GPX4 transcript elucidates the significance of post-transcriptional control in ferroptosis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
FOXM1-activated IGF2BP3 promotes cell malignant phenotypes and M2 macrophage polarization in hepatocellular carcinoma by inhibiting ferroptosis via stabilizing RRM2 mRNA in an m6A-dependent manner.Mol Cell Biochem. 2024 Dec 4. doi: 10.1007/s11010-024-05170-2. Online ahead of print. Mol Cell Biochem. 2024. PMID: 39630361
-
METTL14 plays an oncogenic role in NSCLC by modulating ferroptosis and the m6A modification of GPX4.Arch Physiol Biochem. 2024 Dec;130(6):962-973. doi: 10.1080/13813455.2024.2376813. Epub 2024 Jul 11. Arch Physiol Biochem. 2024. PMID: 38993012
-
Exploring the Mechanism of Ferroptosis Induction by Sappanone A in Cancer: Insights into the Mitochondrial Dysfunction Mediated by NRF2/xCT/GPX4 Axis.Int J Biol Sci. 2024 Sep 23;20(13):5145-5161. doi: 10.7150/ijbs.96748. eCollection 2024. Int J Biol Sci. 2024. PMID: 39430236 Free PMC article.
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
-
The effectiveness of abstinence-based and harm reduction-based interventions in reducing problematic substance use in adults who are experiencing homelessness in high income countries: A systematic review and meta-analysis: A systematic review.Campbell Syst Rev. 2024 Apr 21;20(2):e1396. doi: 10.1002/cl2.1396. eCollection 2024 Jun. Campbell Syst Rev. 2024. PMID: 38645303 Free PMC article. Review.
Cited by
-
IGF2BP3 suppresses ferroptosis in lung adenocarcinoma by m6A-dependent regulation of TFAP2A to transcriptionally activate SLC7A11/GPX4.Mol Cell Biochem. 2024 Jul 18. doi: 10.1007/s11010-024-05068-z. Online ahead of print. Mol Cell Biochem. 2024. PMID: 39026029
-
Deciphering the IGF2BP3-mediated control of ferroptosis: mechanistic insights and therapeutic prospects.Discov Oncol. 2024 Oct 11;15(1):547. doi: 10.1007/s12672-024-01432-z. Discov Oncol. 2024. PMID: 39392532 Free PMC article. Review.
-
Licochalcone A decreases cancer cell proliferation and enhances ferroptosis in acute myeloid leukemia through suppressing the IGF2BP3/MDM2 cascade.Ann Hematol. 2024 Nov;103(11):4511-4524. doi: 10.1007/s00277-024-06003-4. Epub 2024 Sep 12. Ann Hematol. 2024. PMID: 39264435
-
The IGF2BP3/Notch/Jag1 pathway: A key regulator of hepatic stellate cell ferroptosis in liver fibrosis.Clin Transl Med. 2024 Aug;14(8):e1793. doi: 10.1002/ctm2.1793. Clin Transl Med. 2024. PMID: 39113232 Free PMC article.
References
-
- Miller KD, Ostrom QT, Kruchko C, Patil N, Tihan T, Cioffi G, et al. Brain and other central nervous system tumor statistics, 2021. CA: a Cancer J Clin. 2021;71:381–406. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
