

This is a preprint.
Growth-induced collective bending and kinetic trapping of cytoskeletal filaments
- PMID: 38260433
- PMCID: PMC10802417
- DOI: 10.1101/2024.01.09.574885
Growth-induced collective bending and kinetic trapping of cytoskeletal filaments
Update in
-
Growth-induced collective bending and kinetic trapping of cytoskeletal filaments.Cytoskeleton (Hoboken). 2024 Aug;81(8):409-419. doi: 10.1002/cm.21877. Epub 2024 May 22. Cytoskeleton (Hoboken). 2024. PMID: 38775207
Abstract
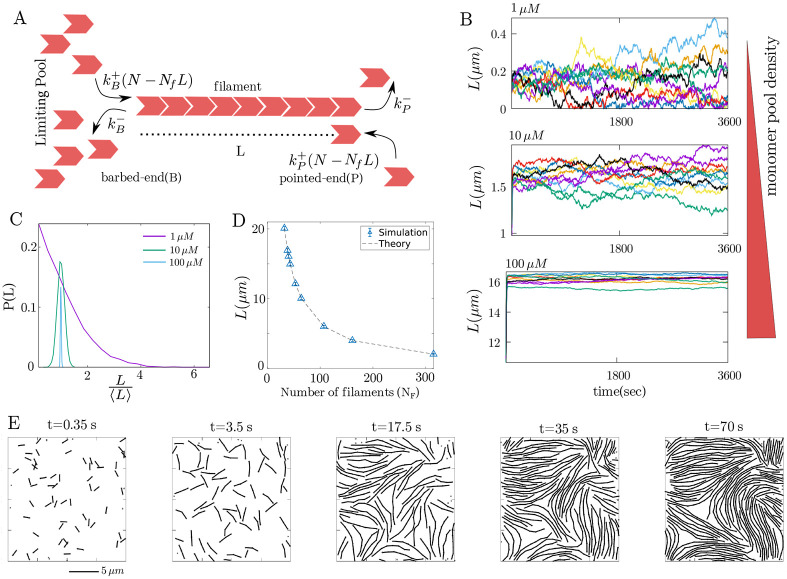
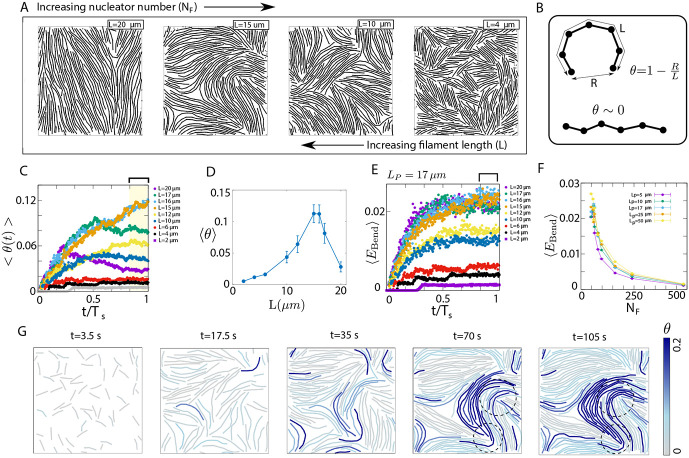
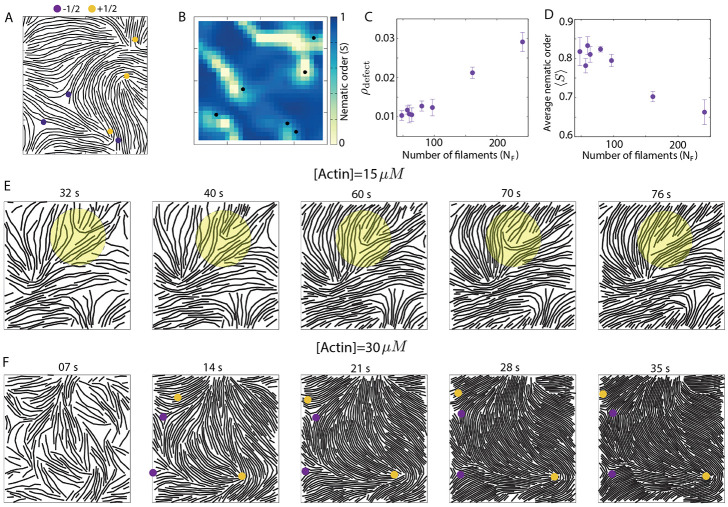
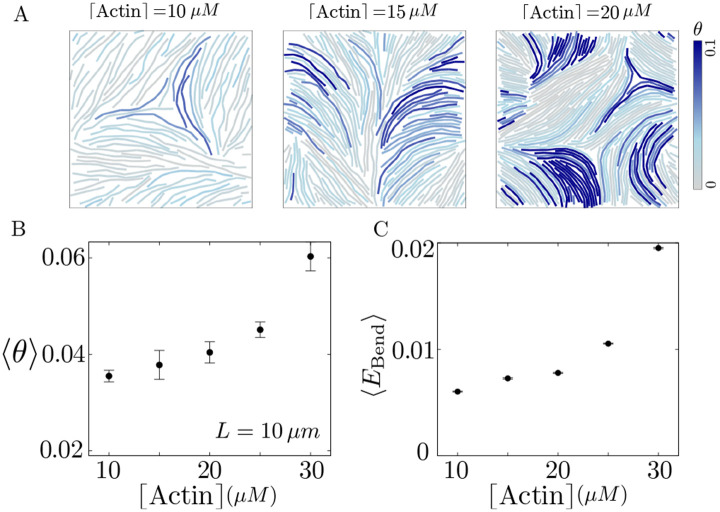
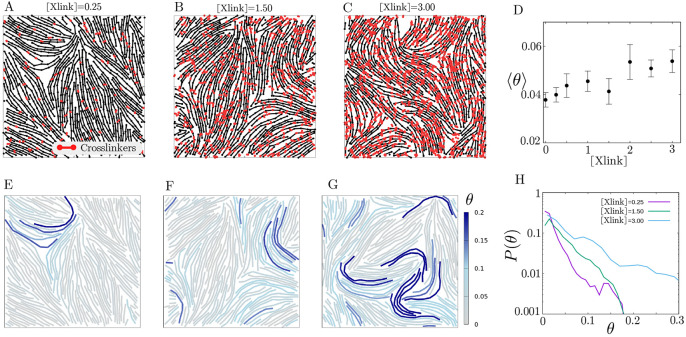
Growth and turnover of actin filaments play a crucial role in the construction and maintenance of actin networks within cells. Actin filament growth occurs within limited space and finite subunit resources in the actin cortex. To understand how filament growth shapes the emergent architecture of actin networks, we developed a minimal agent-based model coupling filament mechanics and growth in a limiting subunit pool. We find that rapid filament growth induces kinetic trapping of highly bent actin filaments. Such collective bending patterns are long-lived, organized around nematic defects, and arises from competition between filament polymerization and bending elasticity. The stability of nematic defects and the extent of kinetic trapping are amplified by an increase in the abundance of the actin pool and by crosslinking the network. These findings suggest that kinetic trapping is a robust consequence of growth in crowded environments, providing a route to program shape memory in actin networks.
Keywords: actin network assembly; agent-based modeling; filament growth; kinetic trapping; topological defects.
Figures
Similar articles
-
Growth-induced collective bending and kinetic trapping of cytoskeletal filaments.Cytoskeleton (Hoboken). 2024 Aug;81(8):409-419. doi: 10.1002/cm.21877. Epub 2024 May 22. Cytoskeleton (Hoboken). 2024. PMID: 38775207
-
Small organic osmolytes accelerate actin filament assembly and stiffen filaments.Cytoskeleton (Hoboken). 2024 Sep 14. doi: 10.1002/cm.21927. Online ahead of print. Cytoskeleton (Hoboken). 2024. PMID: 39276026
-
Origin of twist-bend coupling in actin filaments.Biophys J. 2010 Sep 22;99(6):1852-60. doi: 10.1016/j.bpj.2010.07.009. Biophys J. 2010. PMID: 20858430 Free PMC article.
-
Polymerization and depolymerization of actin with nucleotide states at filament ends.Biophys Rev. 2018 Dec;10(6):1513-1519. doi: 10.1007/s12551-018-0483-7. Epub 2018 Nov 20. Biophys Rev. 2018. PMID: 30460458 Free PMC article. Review.
-
Bending actin filaments: twists of fate.Fac Rev. 2023 Mar 21;12:7. doi: 10.12703/r/12-7. eCollection 2023. Fac Rev. 2023. PMID: 37081903 Free PMC article. Review.
References
-
- Pollard Thomas D and Borisy Gary G. Cellular motility driven by assembly and disassembly of actin filaments. Cell, 112(4):453–465, 2003. - PubMed
-
- Blanchoin Laurent, Boujemaa-Paterski Rajaa, Sykes Cécile, and Plastino Julie. Actin dynamics, architecture, and mechanics in cell motility. Physiological reviews, 94(1):235–263, 2014. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources