

Epitranscriptional m6A modification of rRNA negatively impacts translation and host colonization in Staphylococcus aureus
- PMID: 38252661
- PMCID: PMC10833563
- DOI: 10.1371/journal.ppat.1011968
Epitranscriptional m6A modification of rRNA negatively impacts translation and host colonization in Staphylococcus aureus
Abstract
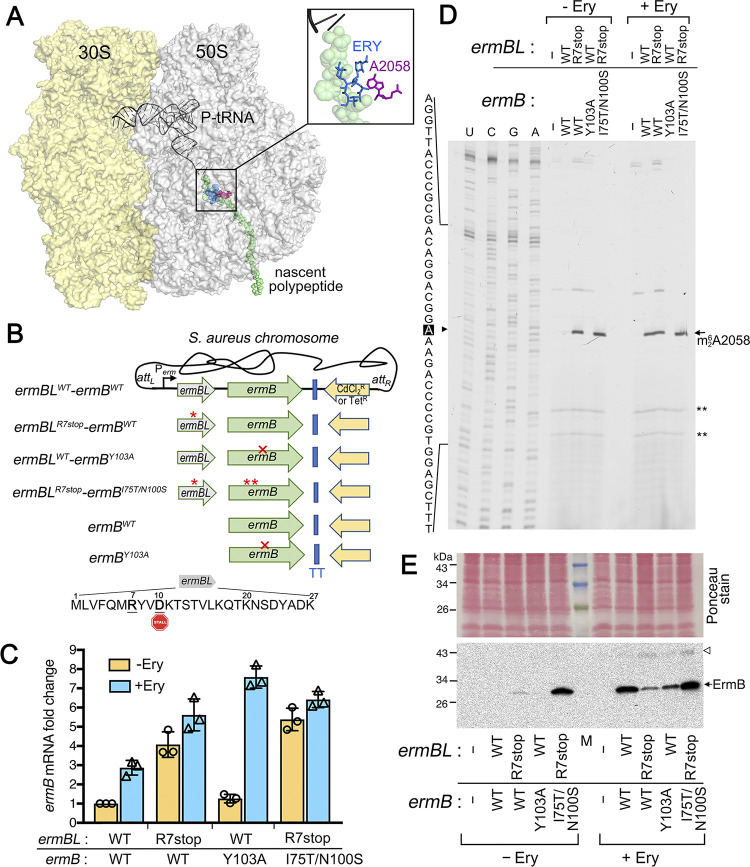
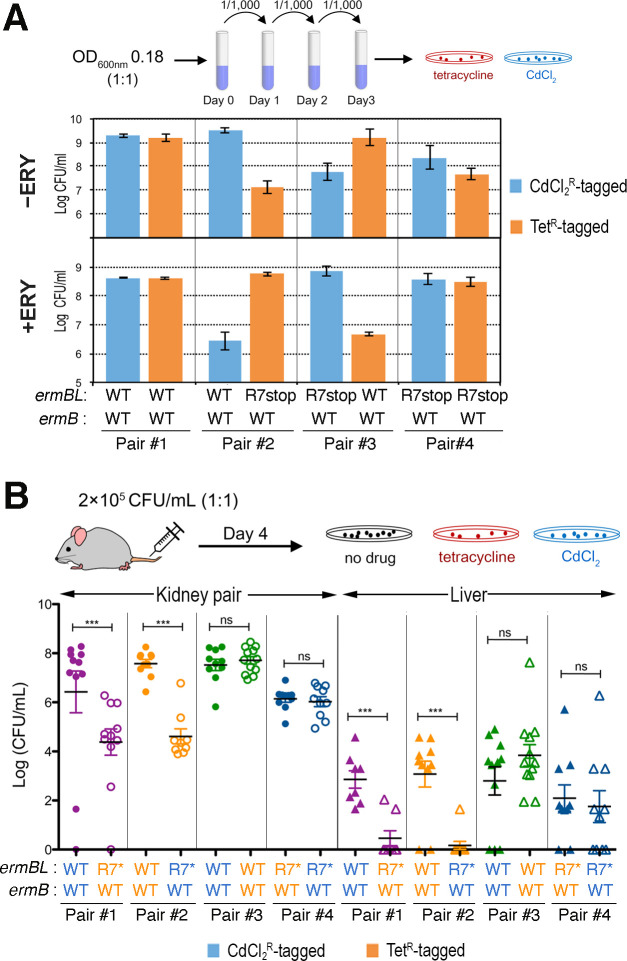
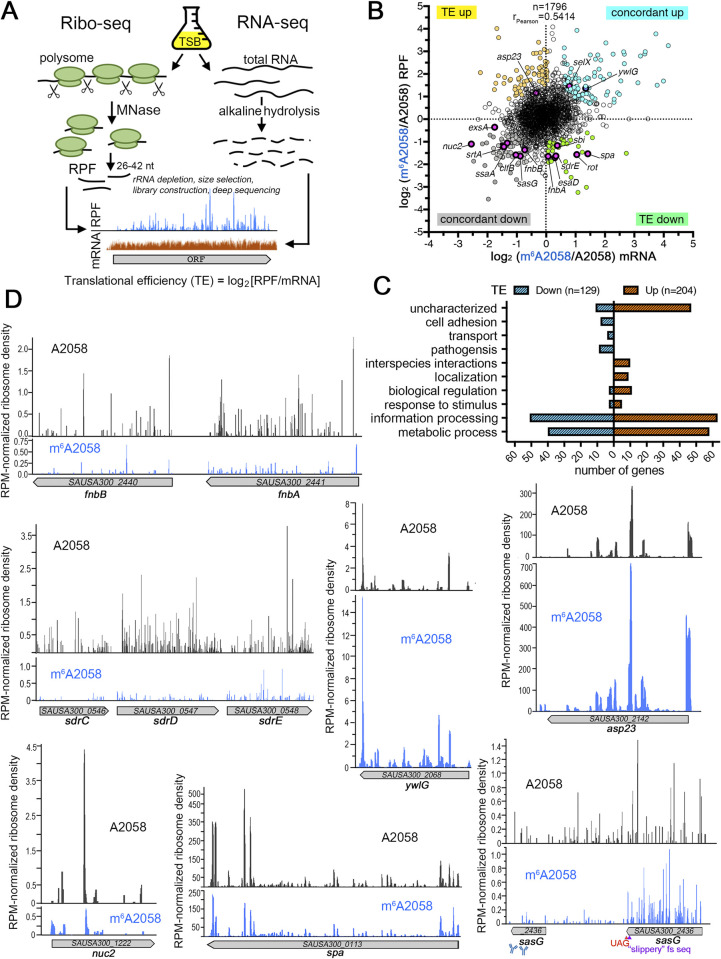
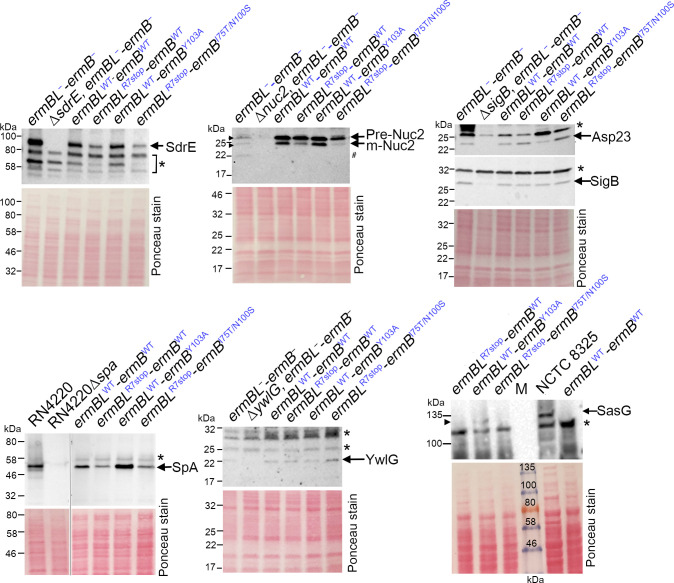
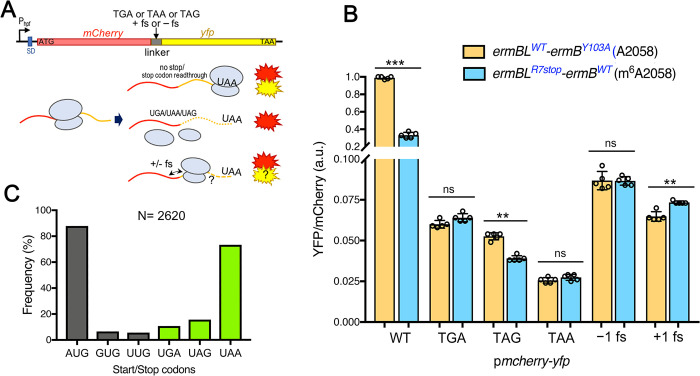
Macrolides, lincosamides, and streptogramin B (MLS) are structurally distinct molecules that are among the safest antibiotics for prophylactic use and for the treatment of bacterial infections. The family of erythromycin resistance methyltransferases (Erm) invariantly install either one or two methyl groups onto the N6,6-adenosine of 2058 nucleotide (m6A2058) of the bacterial 23S rRNA, leading to bacterial cross-resistance to all MLS antibiotics. Despite extensive structural studies on the mechanism of Erm-mediated MLS resistance, how the m6A epitranscriptomic mark affects ribosome function and bacterial physiology is not well understood. Here, we show that Staphylococcus aureus cells harboring m6A2058 ribosomes are outcompeted by cells carrying unmodified ribosomes during infections and are severely impaired in colonization in the absence of an unmodified counterpart. The competitive advantage of m6A2058 ribosomes is manifested only upon antibiotic challenge. Using ribosome profiling (Ribo-Seq) and a dual-fluorescence reporter to measure ribosome occupancy and translational fidelity, we found that specific genes involved in host interactions, metabolism, and information processing are disproportionally deregulated in mRNA translation. This dysregulation is linked to a substantial reduction in translational capacity and fidelity in m6A2058 ribosomes. These findings point to a general "inefficient translation" mechanism of trade-offs associated with multidrug-resistant ribosomes.
Copyright: © 2024 Shields et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Binding of novel macrolide structures to macrolides-lincosamides-streptogramin B-resistant ribosomes inhibits protein synthesis and bacterial growth.Antimicrob Agents Chemother. 1989 Jul;33(7):1058-66. doi: 10.1128/AAC.33.7.1058. Antimicrob Agents Chemother. 1989. PMID: 2506804 Free PMC article.
-
[Study of macrolide, lincosamide, and streptogramin B antibiotics resistance in Staphylococcus aureus].Yakugaku Zasshi. 2000 Apr;120(4):374-86. doi: 10.1248/yakushi1947.120.4_374. Yakugaku Zasshi. 2000. PMID: 10774259 Review. Japanese.
-
Unusual inducible cross resistance to macrolides, lincosamides, and streptogramins B by methylase production in clinical isolates of Staphylococcus aureus.Microb Drug Resist. 2001 Winter;7(4):317-22. doi: 10.1089/10766290152773329. Microb Drug Resist. 2001. PMID: 11822771
-
Distribution of genes encoding resistance to macrolides, lincosamides and streptogramins among clinical staphylococcal isolates in a Turkish university hospital.J Microbiol Immunol Infect. 2010 Dec;43(6):524-9. doi: 10.1016/S1684-1182(10)60081-3. J Microbiol Immunol Infect. 2010. PMID: 21195981
-
Origin and evolution of genes specifying resistance to macrolide, lincosamide and streptogramin antibiotics: data and hypotheses.J Antimicrob Chemother. 1987 Dec;20(6):783-802. doi: 10.1093/jac/20.6.783. J Antimicrob Chemother. 1987. PMID: 3326871 Review.
Cited by
-
Staphylococcal exoribonuclease YhaM destabilizes ribosomes by targeting the mRNA of a hibernation factor.Nucleic Acids Res. 2024 Aug 27;52(15):8998-9013. doi: 10.1093/nar/gkae596. Nucleic Acids Res. 2024. PMID: 38979572 Free PMC article.
-
Machine learning-based classification reveals distinct clusters of non-coding genomic allelic variations associated with Erm-mediated antibiotic resistance.mSystems. 2024 Jul 23;9(7):e0043024. doi: 10.1128/msystems.00430-24. Epub 2024 Jul 2. mSystems. 2024. PMID: 38953319 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
