

Heat tolerance in Drosophila melanogaster is influenced by oxygen conditions and mutations in cell size control pathways
- PMID: 38186282
- PMCID: PMC10772611
- DOI: 10.1098/rstb.2022.0490
Heat tolerance in Drosophila melanogaster is influenced by oxygen conditions and mutations in cell size control pathways
Abstract
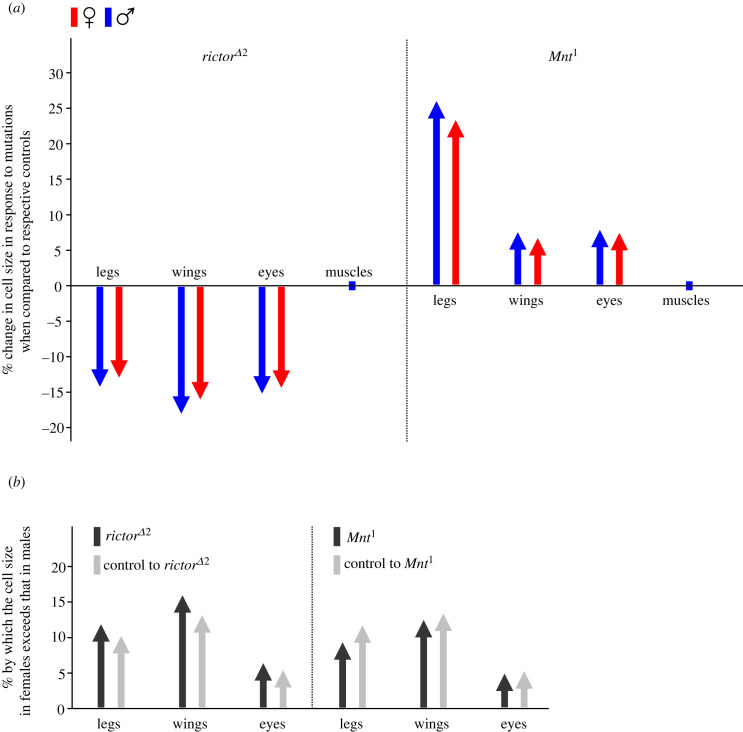
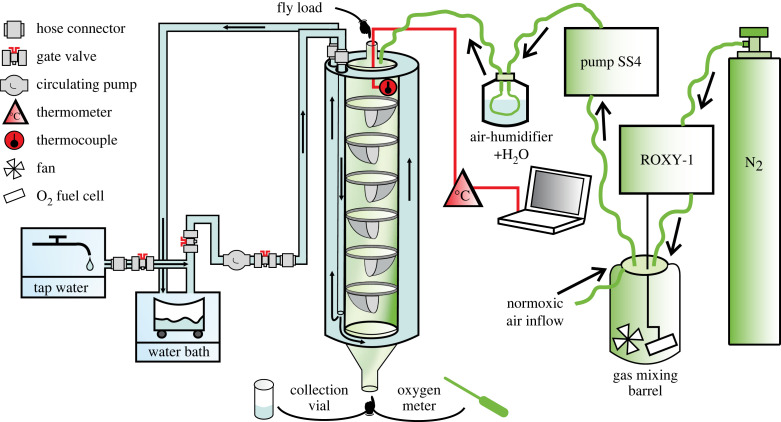
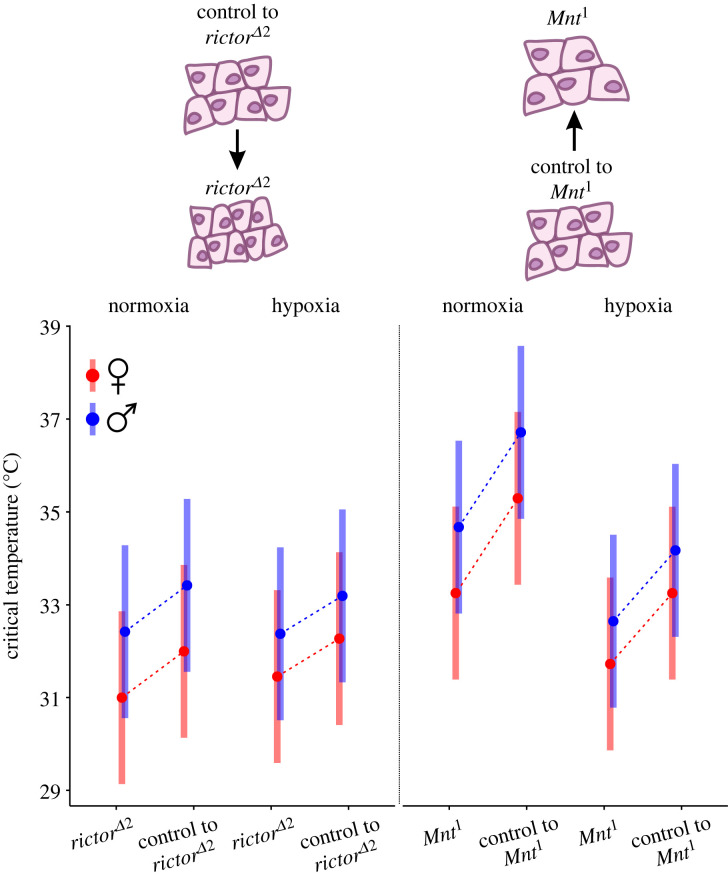
Understanding metabolic performance limitations is key to explaining the past, present and future of life. We investigated whether heat tolerance in actively flying Drosophila melanogaster is modified by individual differences in cell size and the amount of oxygen in the environment. We used two mutants with loss-of-function mutations in cell size control associated with the target of rapamycin (TOR)/insulin pathways, showing reduced (mutant rictorΔ2) or increased (mutant Mnt1) cell size in different body tissues compared to controls. Flies were exposed to a steady increase in temperature under normoxia and hypoxia until they collapsed. The upper critical temperature decreased in response to each mutation type as well as under hypoxia. Females, which have larger cells than males, had lower heat tolerance than males. Altogether, mutations in cell cycle control pathways, differences in cell size and differences in oxygen availability affected heat tolerance, but existing theories on the roles of cell size and tissue oxygenation in metabolic performance can only partially explain our results. A better understanding of how the cellular composition of the body affects metabolism may depend on the development of research models that help separate various interfering physiological parameters from the exclusive influence of cell size. This article is part of the theme issue 'The evolutionary significance of variation in metabolic rates'.
Keywords: Mnt1; TOR; hypoxia; metabolic performance; rictorΔ2; thermal limits.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Systemic changes in cell size throughout the body of Drosophila melanogaster associated with mutations in molecular cell cycle regulators.Sci Rep. 2023 May 9;13(1):7565. doi: 10.1038/s41598-023-34674-y. Sci Rep. 2023. PMID: 37160985 Free PMC article.
-
Effects of developmental plasticity on heat tolerance may be mediated by changes in cell size in Drosophila melanogaster.Insect Sci. 2020 Dec;27(6):1244-1256. doi: 10.1111/1744-7917.12742. Epub 2020 Jan 17. Insect Sci. 2020. PMID: 31829515 Free PMC article.
-
Evolution and development of Drosophila melanogaster under different thermal conditions affected cell sizes and sensitivity to paralyzing hypoxia.J Insect Physiol. 2024 Sep;157:104671. doi: 10.1016/j.jinsphys.2024.104671. Epub 2024 Jul 6. J Insect Physiol. 2024. PMID: 38972633
-
Cellular and developmental adaptations to hypoxia: a Drosophila perspective.Methods Enzymol. 2007;435:123-44. doi: 10.1016/S0076-6879(07)35007-6. Methods Enzymol. 2007. PMID: 17998052 Review.
-
Effects of oxygen on growth and size: synthesis of molecular, organismal, and evolutionary studies with Drosophila melanogaster.Annu Rev Physiol. 2011;73:95-113. doi: 10.1146/annurev-physiol-012110-142155. Annu Rev Physiol. 2011. PMID: 20936942 Review.
Cited by
-
Consequences of the cost of living: is variation in metabolic rate evolutionarily significant?Philos Trans R Soc Lond B Biol Sci. 2024 Feb 26;379(1896):20220498. doi: 10.1098/rstb.2022.0498. Epub 2024 Jan 8. Philos Trans R Soc Lond B Biol Sci. 2024. PMID: 38186277 Free PMC article. No abstract available.
References
-
- Clarke A. 2014. The thermal limits to life on Earth. Int. J. Astrobiol. 13, 141-154. (10.1017/S1473550413000438) - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
