

The Roles of eIF4G2 in Leaky Scanning and Reinitiation on the Human Dual-Coding POLG mRNA
- PMID: 38138978
- PMCID: PMC10742948
- DOI: 10.3390/ijms242417149
The Roles of eIF4G2 in Leaky Scanning and Reinitiation on the Human Dual-Coding POLG mRNA
Abstract
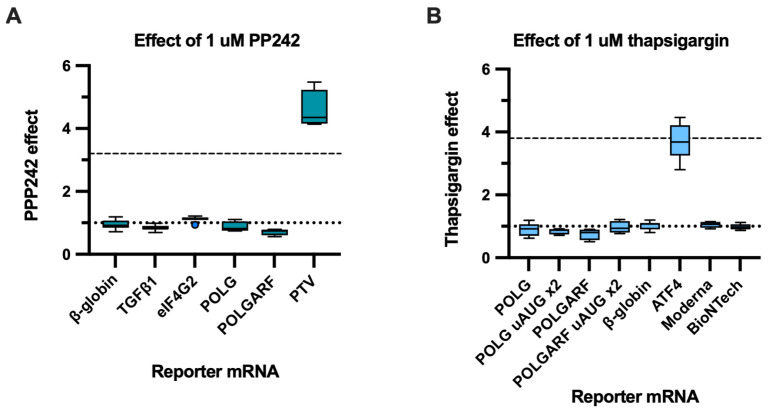
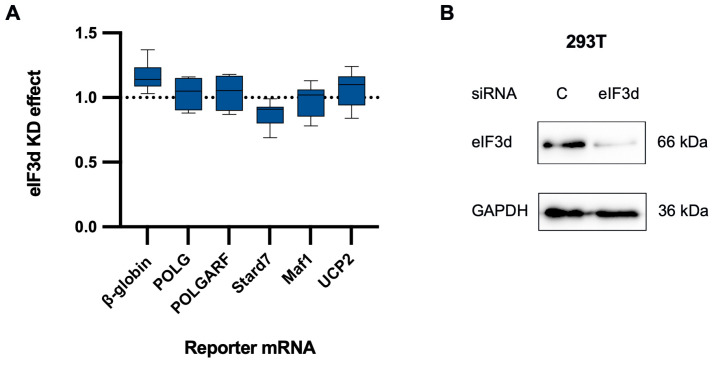
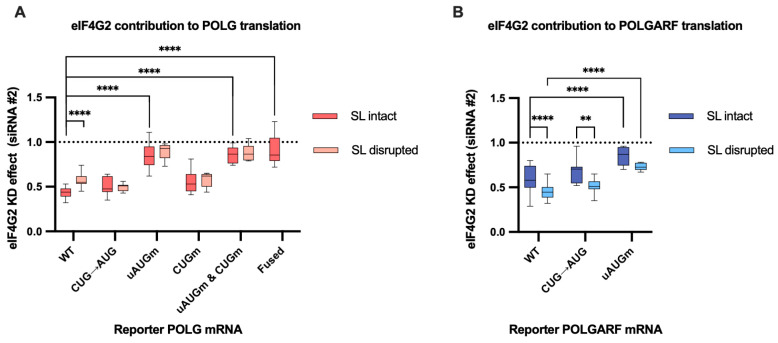
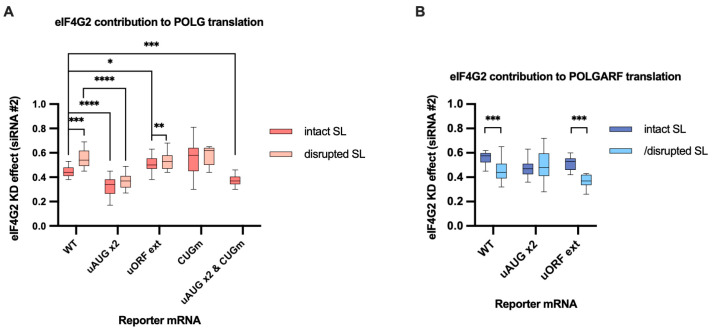
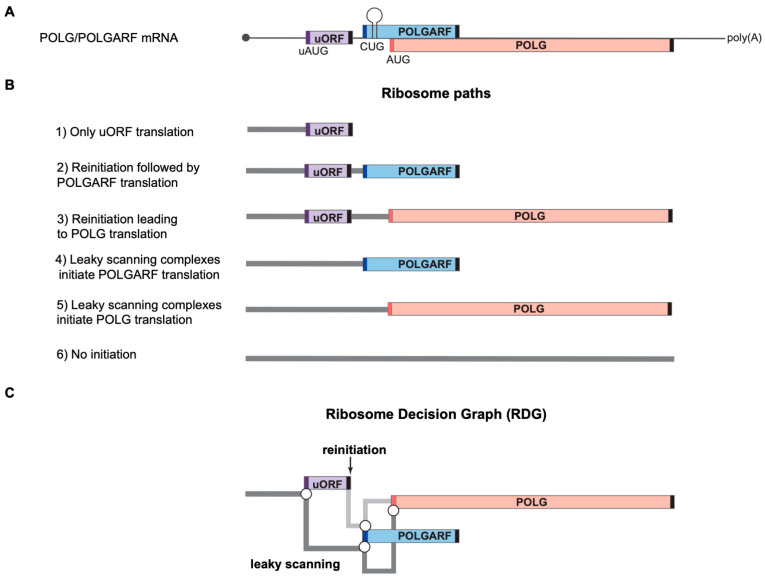
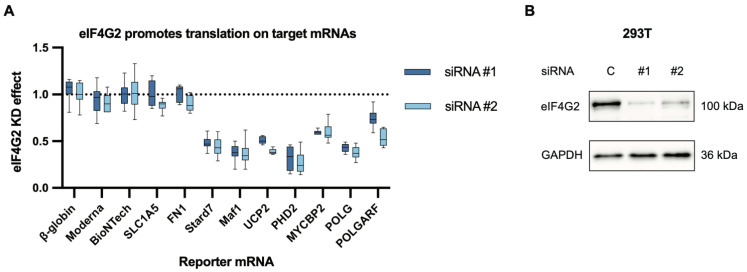
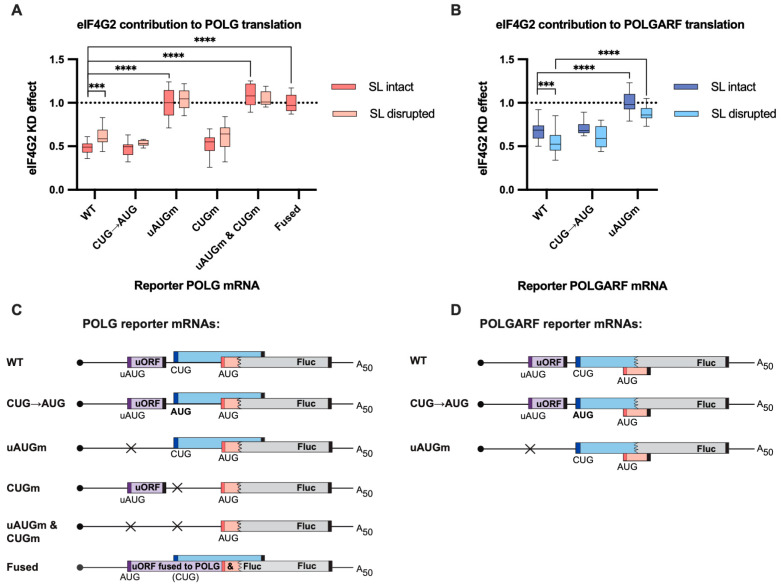
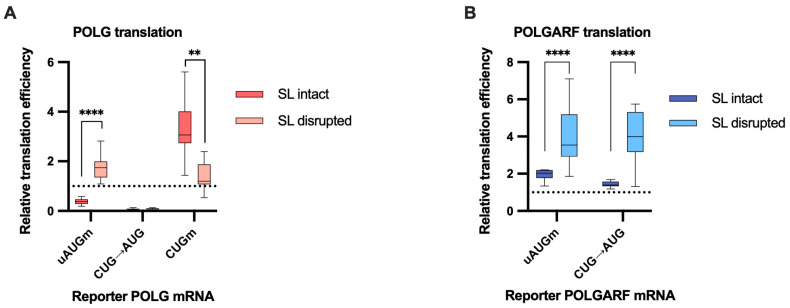
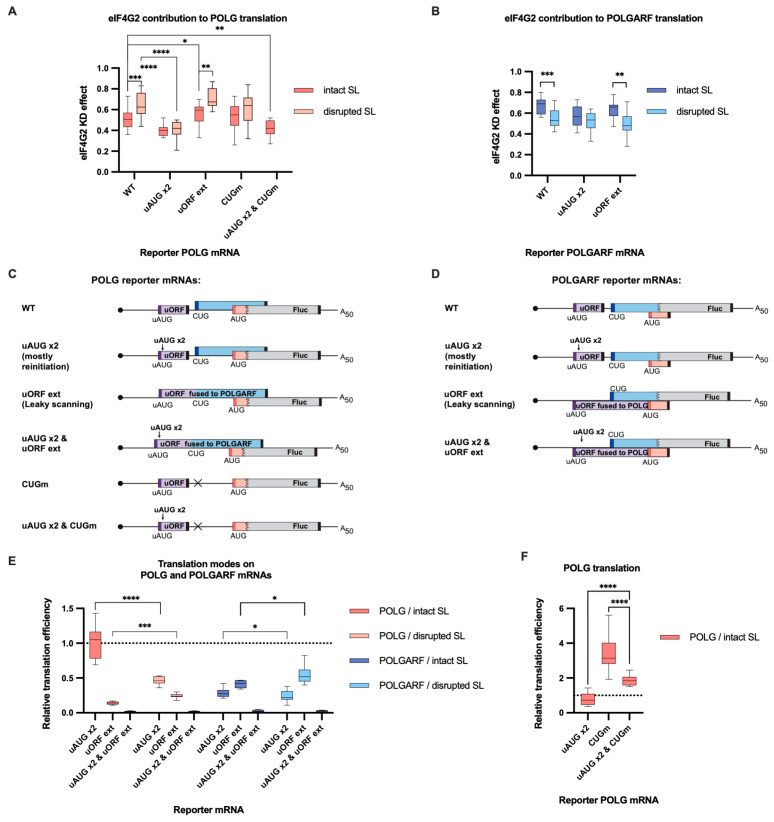
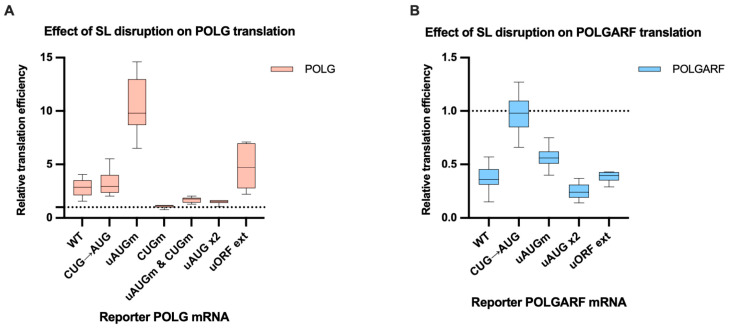
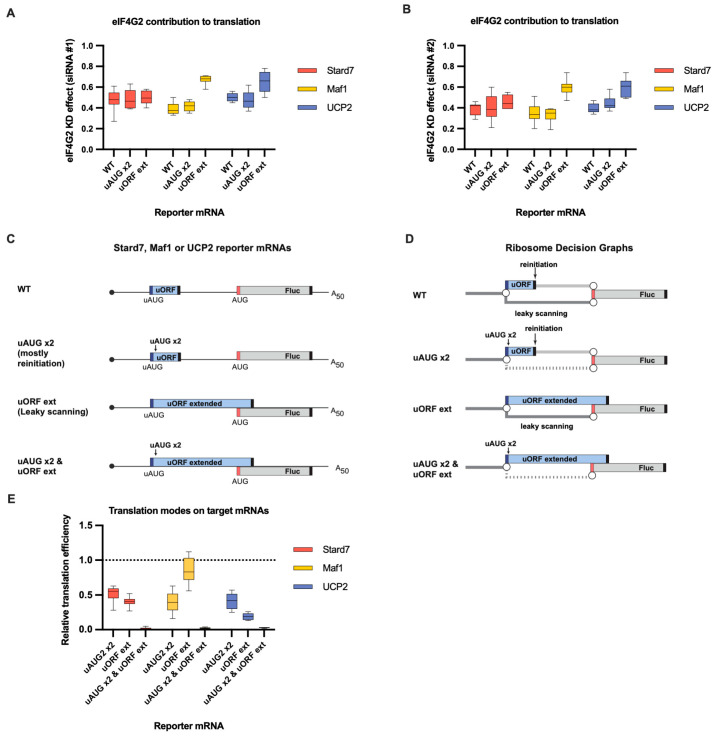
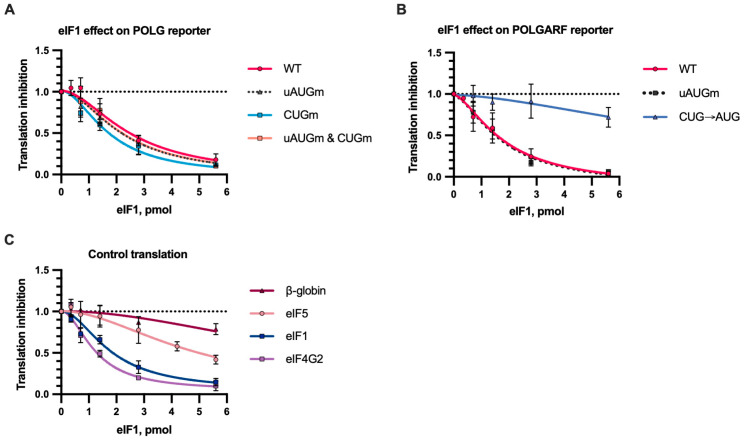
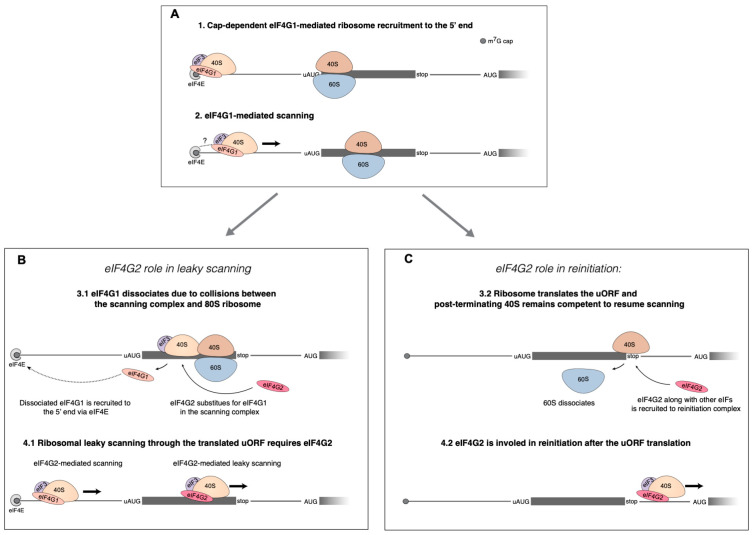
Upstream open reading frames (uORFs) are a frequent feature of eukaryotic mRNAs. Upstream ORFs govern main ORF translation in a variety of ways, but, in a nutshell, they either filter out scanning ribosomes or allow downstream translation initiation via leaky scanning or reinitiation. Previous reports concurred that eIF4G2, a long-known but insufficiently studied eIF4G1 homologue, can rescue the downstream translation, but disagreed on whether it is leaky scanning or reinitiation that eIF4G2 promotes. Here, we investigated a unique human mRNA that encodes two highly conserved proteins (POLGARF with unknown function and POLG, the catalytic subunit of the mitochondrial DNA polymerase) in overlapping reading frames downstream of a regulatory uORF. We show that the uORF renders the translation of both POLGARF and POLG mRNAs reliant on eIF4G2. Mechanistically, eIF4G2 enhances both leaky scanning and reinitiation, and it appears that ribosomes can acquire eIF4G2 during the early steps of reinitiation. This emphasizes the role of eIF4G2 as a multifunctional scanning guardian that replaces eIF4G1 to facilitate ribosome movement but not ribosome attachment to an mRNA.
Keywords: 40S; AUG selection; MYCBP2; PHD2; cap-dependent translation; mitochondrial disfunction; non-AUG translation; polyglutamine; ribosome collision; translation reinitiation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Ribosomal leaky scanning through a translated uORF requires eIF4G2.Nucleic Acids Res. 2022 Jan 25;50(2):1111-1127. doi: 10.1093/nar/gkab1286. Nucleic Acids Res. 2022. PMID: 35018467 Free PMC article.
-
Specific mechanisms of translation initiation in higher eukaryotes: the eIF4G2 story.RNA. 2023 Mar;29(3):282-299. doi: 10.1261/rna.079462.122. Epub 2022 Dec 14. RNA. 2023. PMID: 36517212 Free PMC article. Review.
-
Unusually efficient CUG initiation of an overlapping reading frame in POLG mRNA yields novel protein POLGARF.Proc Natl Acad Sci U S A. 2020 Oct 6;117(40):24936-24946. doi: 10.1073/pnas.2001433117. Epub 2020 Sep 21. Proc Natl Acad Sci U S A. 2020. PMID: 32958672 Free PMC article.
-
5'-untranslated regions with multiple upstream AUG codons can support low-level translation via leaky scanning and reinitiation.Nucleic Acids Res. 2004 Feb 27;32(4):1382-91. doi: 10.1093/nar/gkh305. Print 2004. Nucleic Acids Res. 2004. PMID: 14990743 Free PMC article.
-
Does eIF3 promote reinitiation after translation of short upstream ORFs also in mammalian cells?RNA Biol. 2017 Dec 2;14(12):1660-1667. doi: 10.1080/15476286.2017.1353863. Epub 2017 Sep 15. RNA Biol. 2017. PMID: 28745933 Free PMC article. Review.
Cited by
-
Evidence for widespread translation of 5' untranslated regions.Nucleic Acids Res. 2024 Aug 12;52(14):8112-8126. doi: 10.1093/nar/gkae571. Nucleic Acids Res. 2024. PMID: 38953162 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
