

Serine-129 phosphorylation of α-synuclein is an activity-dependent trigger for physiologic protein-protein interactions and synaptic function
- PMID: 38128479
- PMCID: PMC10766085
- DOI: 10.1016/j.neuron.2023.11.020
Serine-129 phosphorylation of α-synuclein is an activity-dependent trigger for physiologic protein-protein interactions and synaptic function
Abstract
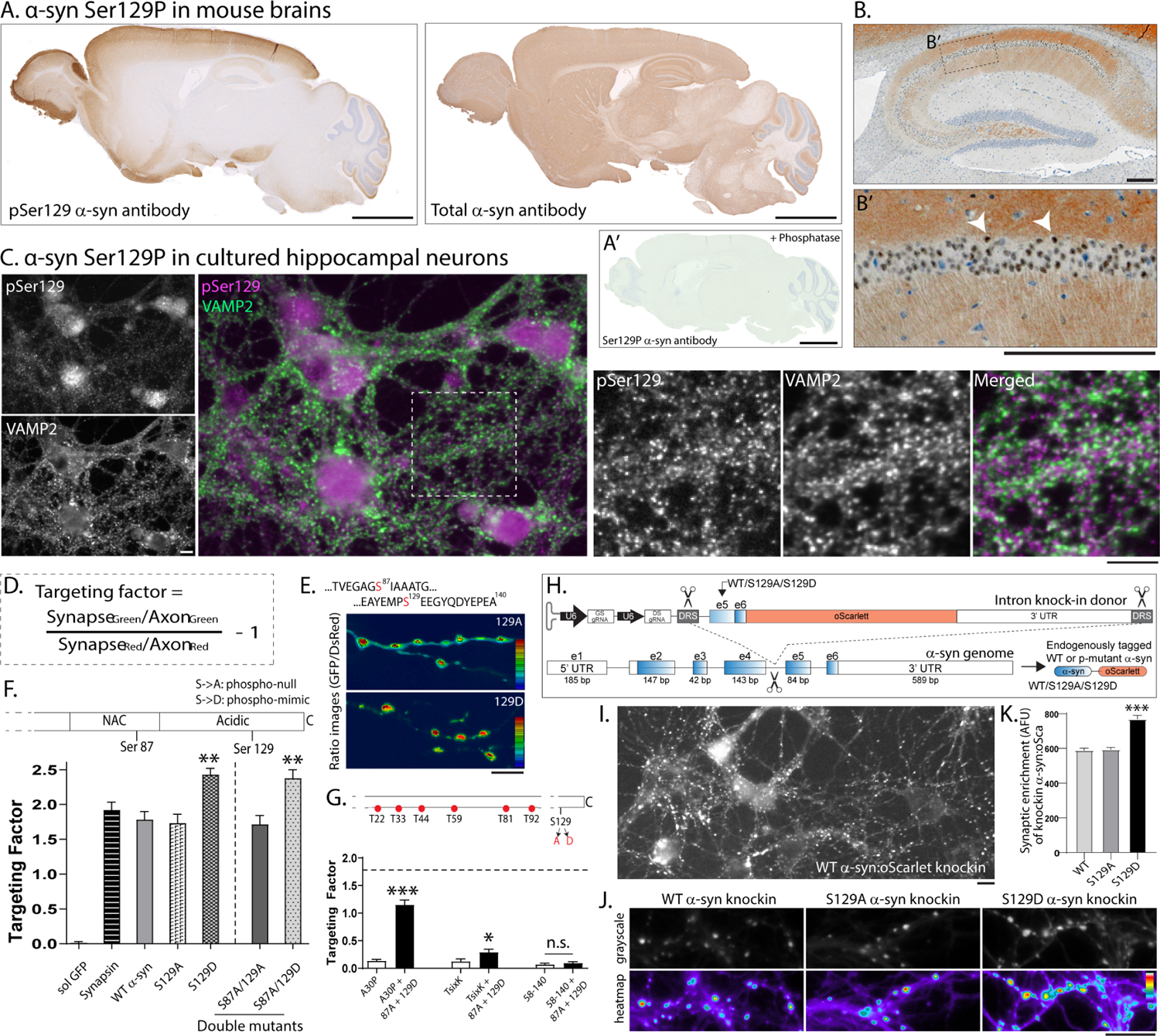
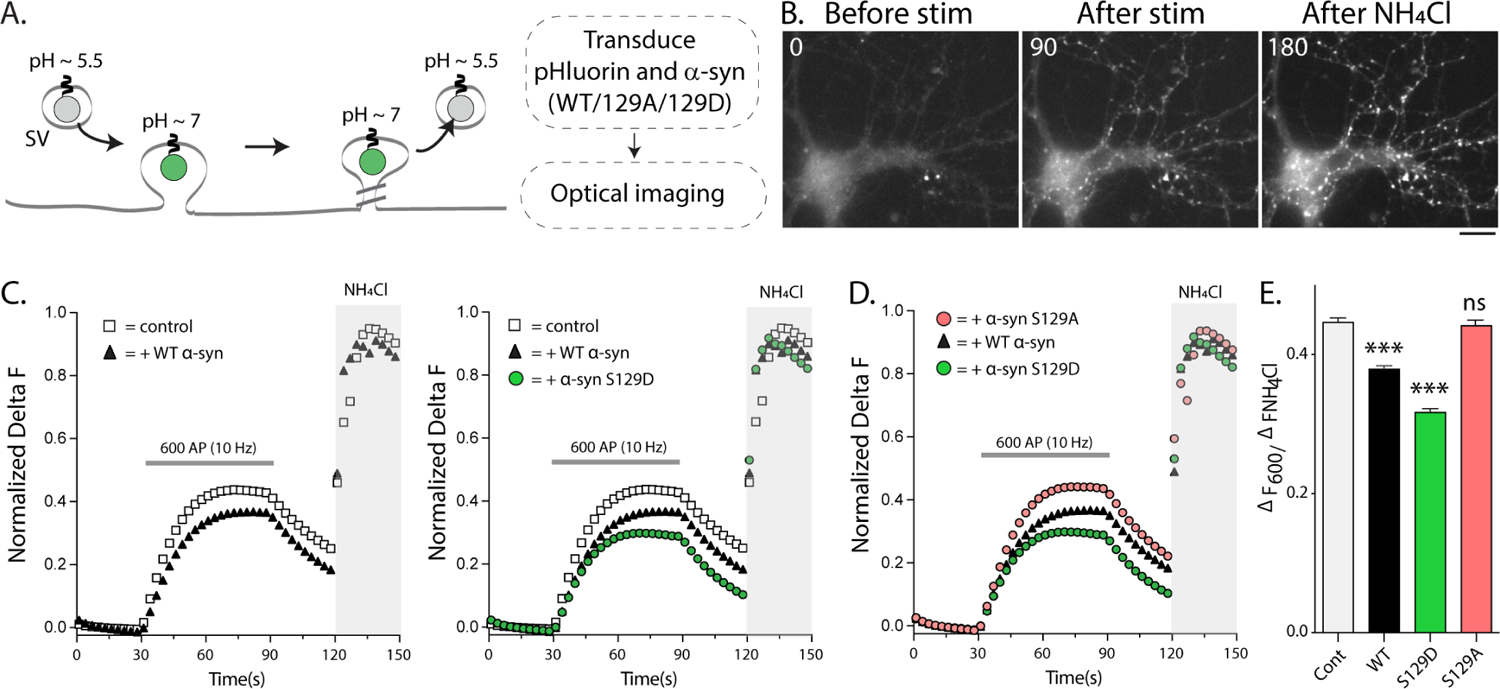
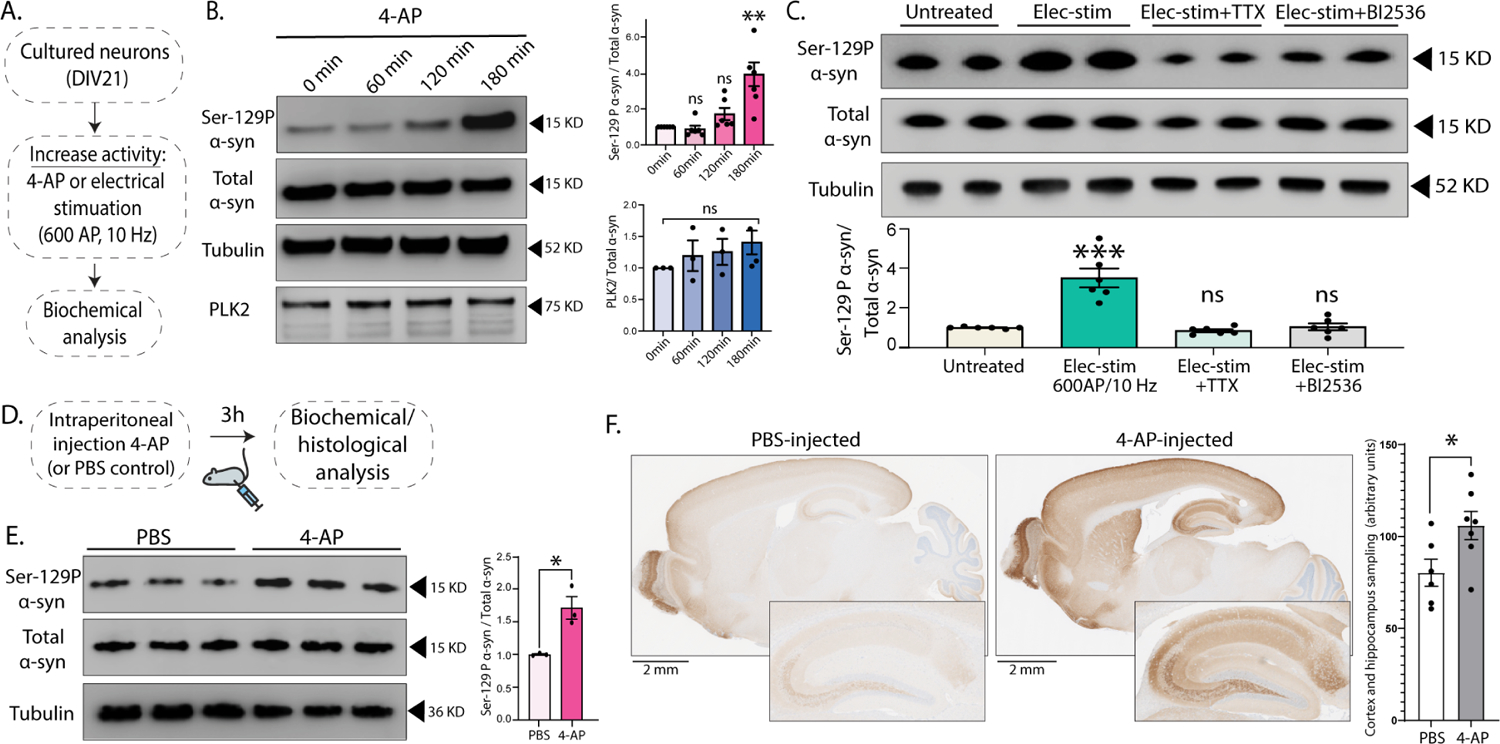
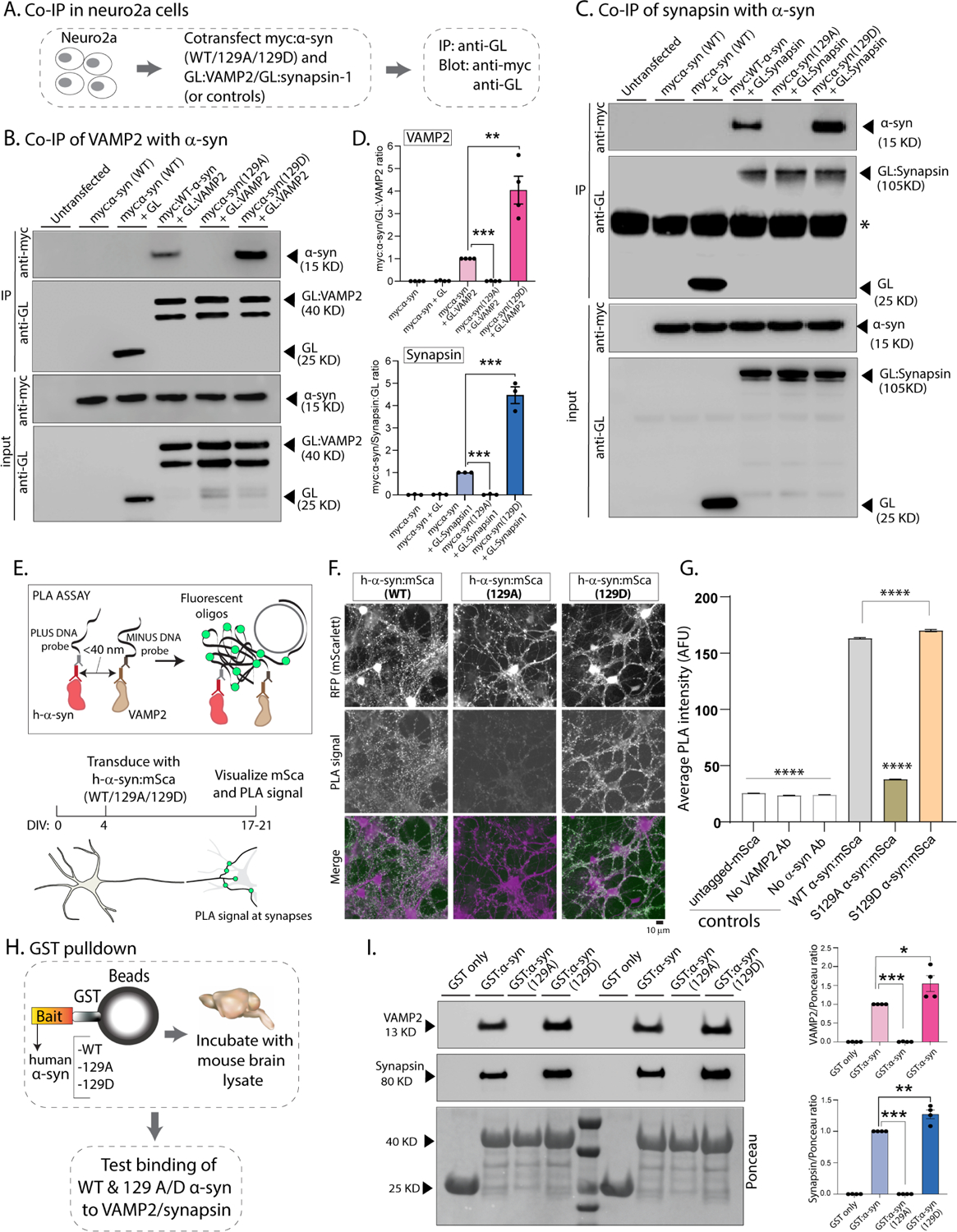
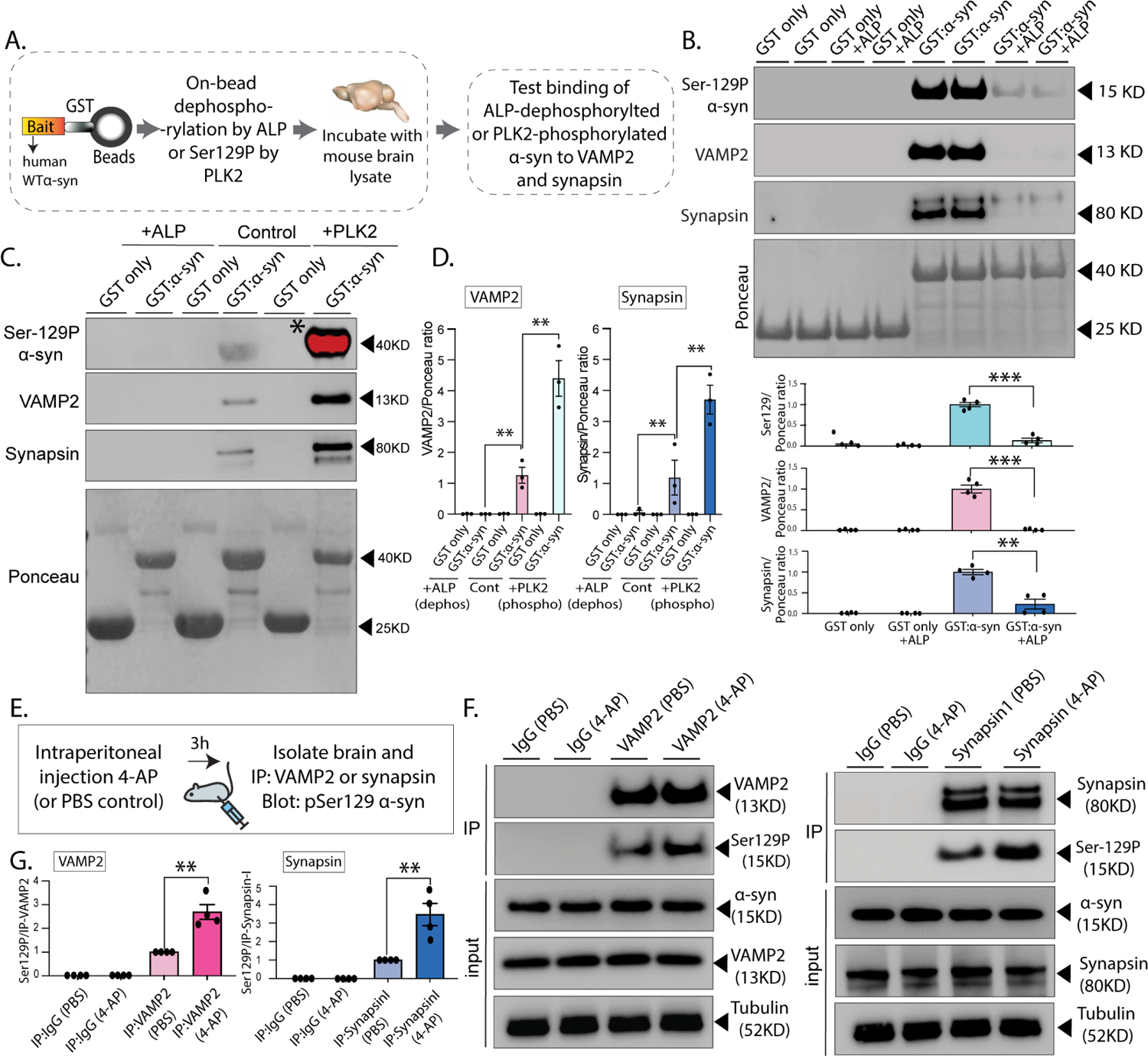
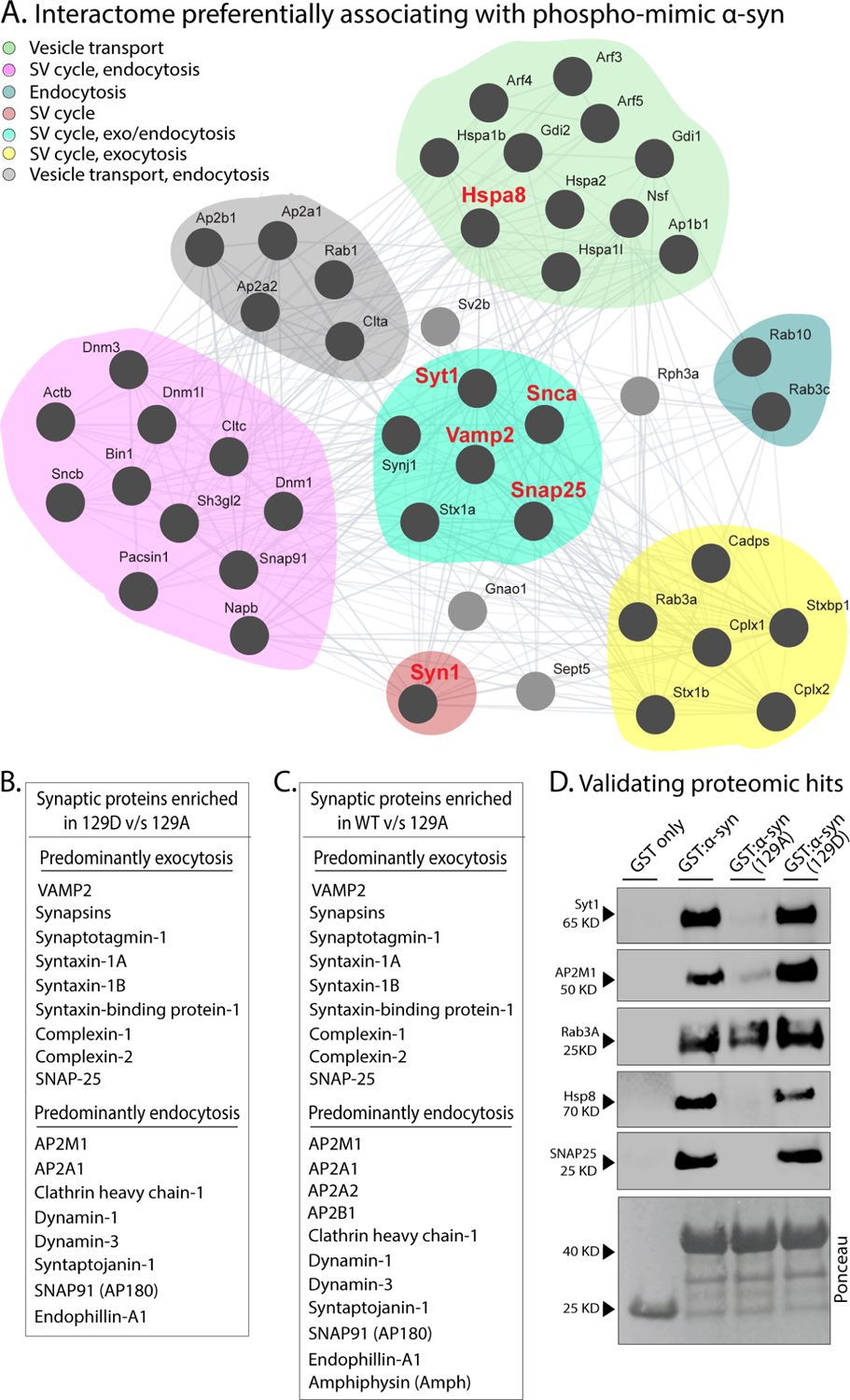
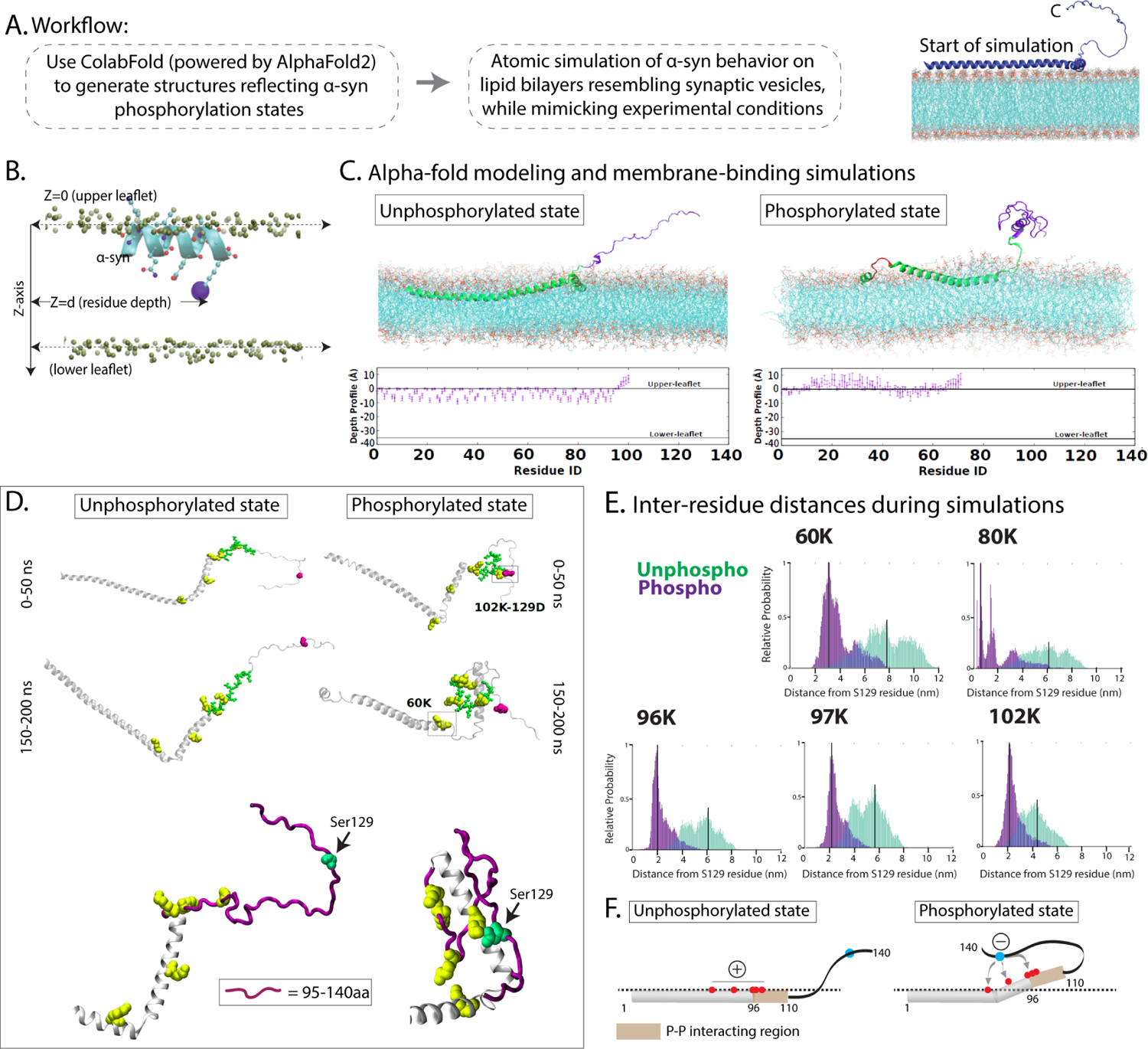
Phosphorylation of α-synuclein at the serine-129 site (α-syn Ser129P) is an established pathologic hallmark of synucleinopathies and a therapeutic target. In physiologic states, only a fraction of α-syn is phosphorylated at this site, and most studies have focused on the pathologic roles of this post-translational modification. We found that unlike wild-type (WT) α-syn, which is widely expressed throughout the brain, the overall pattern of α-syn Ser129P is restricted, suggesting intrinsic regulation. Surprisingly, preventing Ser129P blocked activity-dependent synaptic attenuation by α-syn-thought to reflect its normal function. Exploring mechanisms, we found that neuronal activity augments Ser129P, which is a trigger for protein-protein interactions that are necessary for mediating α-syn function at the synapse. AlphaFold2-driven modeling and membrane-binding simulations suggest a scenario where Ser129P induces conformational changes that facilitate interactions with binding partners. Our experiments offer a new conceptual platform for investigating the role of Ser129 in synucleinopathies, with implications for drug development.
Keywords: AlphaFold; CRISPR; alanine mutations; endogenous tagging; neuronal activity; pHluorin; proximity-ligation assay; serine phosphorylation; synapses; α-synuclein.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures

Similar articles
-
Brain-derived and in vitro-seeded alpha-synuclein fibrils exhibit distinct biophysical profiles.Elife. 2024 Nov 25;13:RP92775. doi: 10.7554/eLife.92775. Elife. 2024. PMID: 39584804 Free PMC article.
-
Defining the optimum strategy for identifying adults and children with coeliac disease: systematic review and economic modelling.Health Technol Assess. 2022 Oct;26(44):1-310. doi: 10.3310/ZUCE8371. Health Technol Assess. 2022. PMID: 36321689 Free PMC article.
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
-
Dynamic Field Theory of Executive Function: Identifying Early Neurocognitive Markers.Monogr Soc Res Child Dev. 2024 Dec;89(3):7-109. doi: 10.1111/mono.12478. Monogr Soc Res Child Dev. 2024. PMID: 39628288 Free PMC article.
-
Impact of residual disease as a prognostic factor for survival in women with advanced epithelial ovarian cancer after primary surgery.Cochrane Database Syst Rev. 2022 Sep 26;9(9):CD015048. doi: 10.1002/14651858.CD015048.pub2. Cochrane Database Syst Rev. 2022. PMID: 36161421 Free PMC article. Review.
Cited by
-
Excess phosphoserine-129 α-synuclein induces synaptic vesicle trafficking and declustering defects at a vertebrate synapse.Mol Biol Cell. 2024 Jan 1;35(1):ar10. doi: 10.1091/mbc.E23-07-0269. Epub 2023 Nov 22. Mol Biol Cell. 2024. PMID: 37991902 Free PMC article.
-
Biochemical Fractionation of Human α-Synuclein in a Drosophila Model of Synucleinopathies.Int J Mol Sci. 2024 Mar 25;25(7):3643. doi: 10.3390/ijms25073643. Int J Mol Sci. 2024. PMID: 38612454 Free PMC article.
-
A-Syn(ful) MAM: A Fresh Perspective on a Converging Domain in Parkinson's Disease.Int J Mol Sci. 2024 Jun 13;25(12):6525. doi: 10.3390/ijms25126525. Int J Mol Sci. 2024. PMID: 38928232 Free PMC article. Review.
-
Motor deficits and brain pathology in the Parkinson's disease mouse model hA53Ttg.Front Neurosci. 2024 Sep 20;18:1462041. doi: 10.3389/fnins.2024.1462041. eCollection 2024. Front Neurosci. 2024. PMID: 39371610 Free PMC article.
-
α-Synuclein serine129 phosphorylation - the physiology of pathology.Mol Neurodegener. 2023 Nov 13;18(1):84. doi: 10.1186/s13024-023-00680-x. Mol Neurodegener. 2023. PMID: 37953316 Free PMC article. No abstract available.
References
-
- Anderson JP, Walker DE, Goldstein JM, de Laat R, Banducci K, Caccavello RJ, Barbour R, Huang J, Kling K, Lee M, et al. (2006). Phosphorylation of Ser-129 is the dominant pathological modification of alpha-synuclein in familial and sporadic Lewy body disease. J Biol Chem 281, 29739–29752. M600933200 [pii]10.1074/jbc.M600933200. - DOI - PubMed
-
- Fujiwara H, Hasegawa M, Dohmae N, Kawashima A, Masliah E, Goldberg MS, Shen J, Takio K, and Iwatsubo T (2002). alpha-Synuclein is phosphorylated in synucleinopathy lesions. Nat.Cell Biol 4, 160–164. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
