

The competitive landscape of the dsRNA world
- PMID: 38118451
- PMCID: PMC10843539
- DOI: 10.1016/j.molcel.2023.11.033
The competitive landscape of the dsRNA world
Abstract
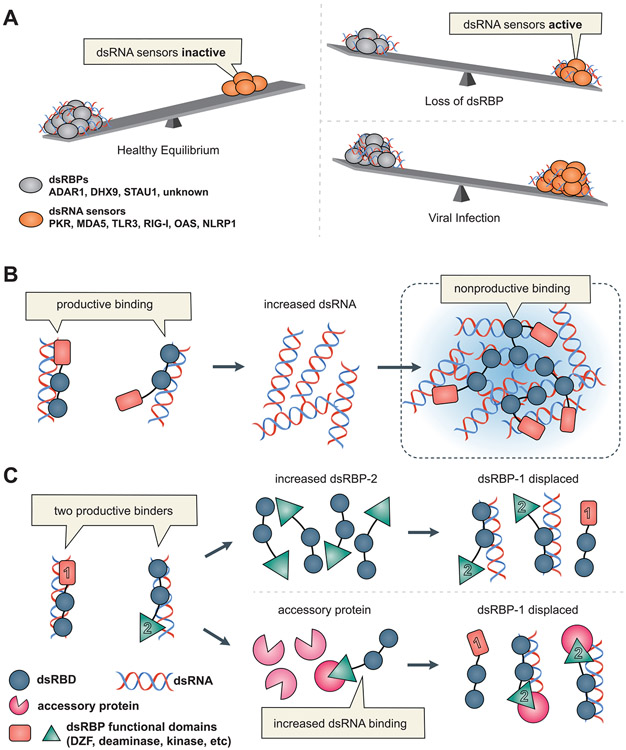
The ability to sense and respond to infection is essential for life. Viral infection produces double-stranded RNAs (dsRNAs) that are sensed by proteins that recognize the structure of dsRNA. This structure-based recognition of viral dsRNA allows dsRNA sensors to recognize infection by many viruses, but it comes at a cost-the dsRNA sensors cannot always distinguish between "self" and "nonself" dsRNAs. "Self" RNAs often contain dsRNA regions, and not surprisingly, mechanisms have evolved to prevent aberrant activation of dsRNA sensors by "self" RNA. Here, we review current knowledge about the life of endogenous dsRNAs in mammals-the biosynthesis and processing of dsRNAs, the proteins they encounter, and their ultimate degradation. We highlight mechanisms that evolved to prevent aberrant dsRNA sensor activation and the importance of competition in the regulation of dsRNA sensors and other dsRNA-binding proteins.
Keywords: ADAR; DHX9; MDA5; PKR; RIG-I; antiviral; dsRNA-binding proteins; innate immunity; interferon.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures

Similar articles
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
-
The double-stranded RNA-dependent protein kinase PKR negatively regulates the protein expression of IFN-β induced by RIG-I signaling.FASEB J. 2023 Feb;37(2):e22780. doi: 10.1096/fj.202201520RR. FASEB J. 2023. PMID: 36651716
-
The vaccinia virus E3L dsRNA binding protein detects distinct production patterns of exogenous and endogenous dsRNA.bioRxiv [Preprint]. 2023 Sep 21:2023.09.21.557600. doi: 10.1101/2023.09.21.557600. bioRxiv. 2023. PMID: 37790463 Free PMC article. Preprint.
-
Hepatitis C Virus Infection Is Inhibited by a Noncanonical Antiviral Signaling Pathway Targeted by NS3-NS4A.J Virol. 2019 Nov 13;93(23):e00725-19. doi: 10.1128/JVI.00725-19. Print 2019 Dec 1. J Virol. 2019. PMID: 31534039 Free PMC article.
-
DEAD-box RNA helicases: The driving forces behind RNA metabolism at the crossroad of viral replication and antiviral innate immunity.Virus Res. 2021 Apr 15;296:198352. doi: 10.1016/j.virusres.2021.198352. Epub 2021 Feb 25. Virus Res. 2021. PMID: 33640359 Review.
Cited by
-
Natural antisense transcripts as versatile regulators of gene expression.Nat Rev Genet. 2024 Oct;25(10):730-744. doi: 10.1038/s41576-024-00723-z. Epub 2024 Apr 17. Nat Rev Genet. 2024. PMID: 38632496 Review.
-
The Sixth Sense: Self-nucleic acid sensing in the brain.Adv Immunol. 2024;161:53-83. doi: 10.1016/bs.ai.2024.03.001. Epub 2024 May 3. Adv Immunol. 2024. PMID: 38763702 Free PMC article. Review.
-
Adenosine deaminases that act on RNA, then and now.RNA. 2024 Apr 16;30(5):521-529. doi: 10.1261/rna.079990.124. RNA. 2024. PMID: 38531651 Free PMC article.
-
Activation of PKR by a short-hairpin RNA.Sci Rep. 2024 Oct 9;14(1):23533. doi: 10.1038/s41598-024-74477-3. Sci Rep. 2024. PMID: 39384561 Free PMC article.
-
Activation of PKR by a short-hairpin RNA.bioRxiv [Preprint]. 2024 May 10:2024.05.08.592371. doi: 10.1101/2024.05.08.592371. bioRxiv. 2024. Update in: Sci Rep. 2024 Oct 9;14(1):23533. doi: 10.1038/s41598-024-74477-3. PMID: 38766230 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
