

Active condensation of filaments under spatial confinement
- PMID: 38116396
- PMCID: PMC10730113
- DOI: 10.3389/fphy.2022.897255
Active condensation of filaments under spatial confinement
Abstract
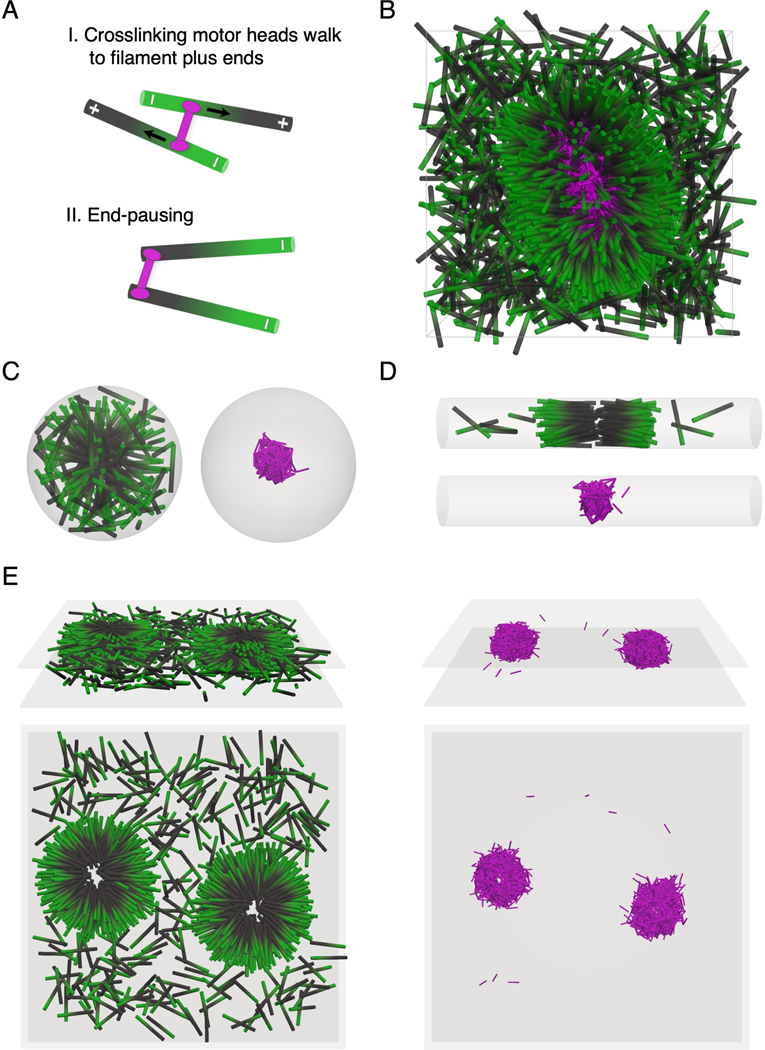
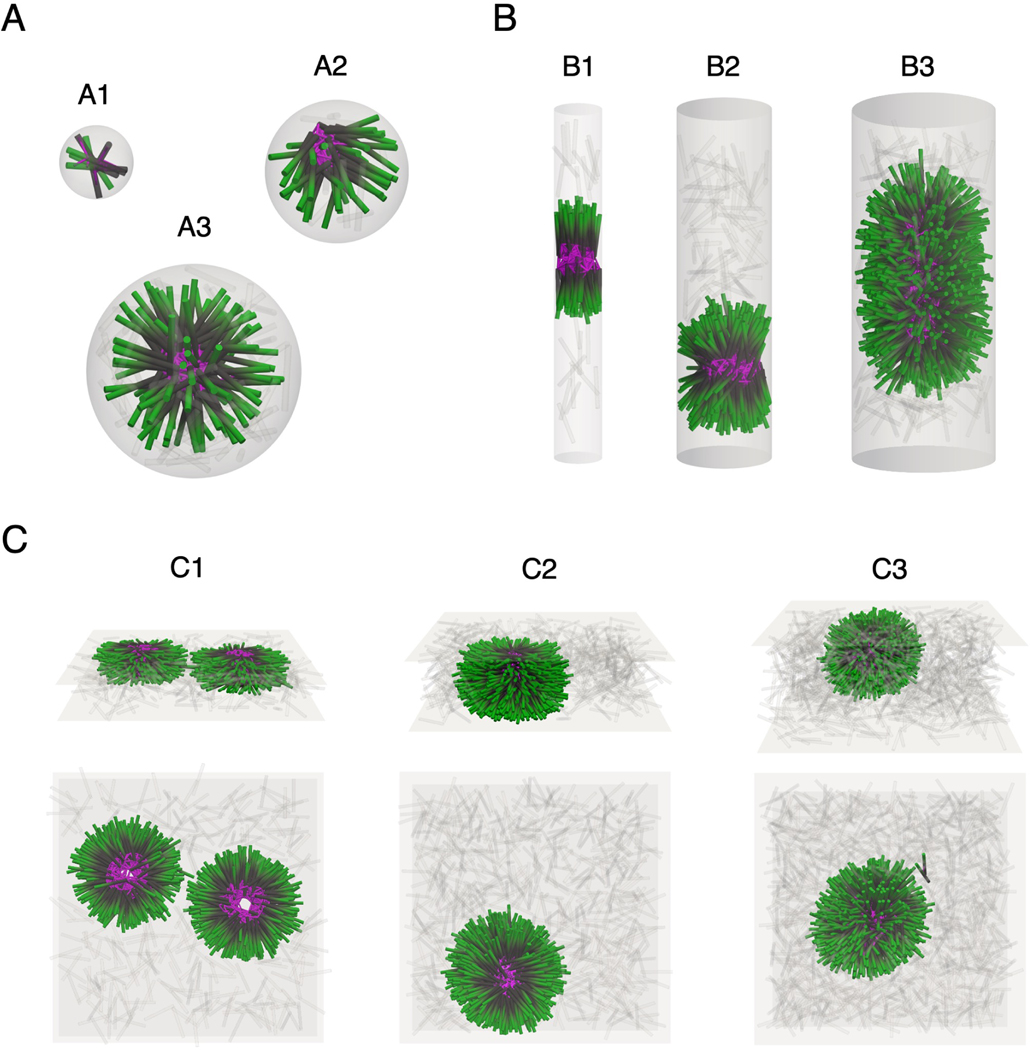
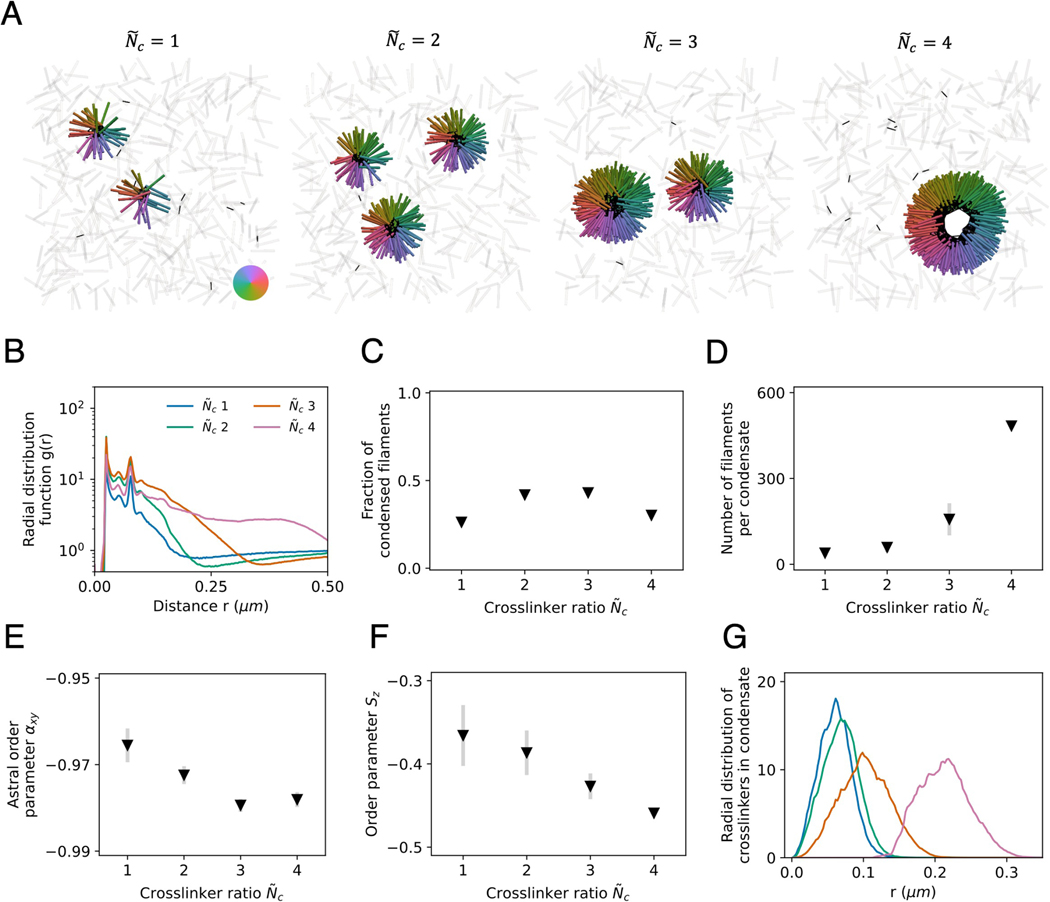
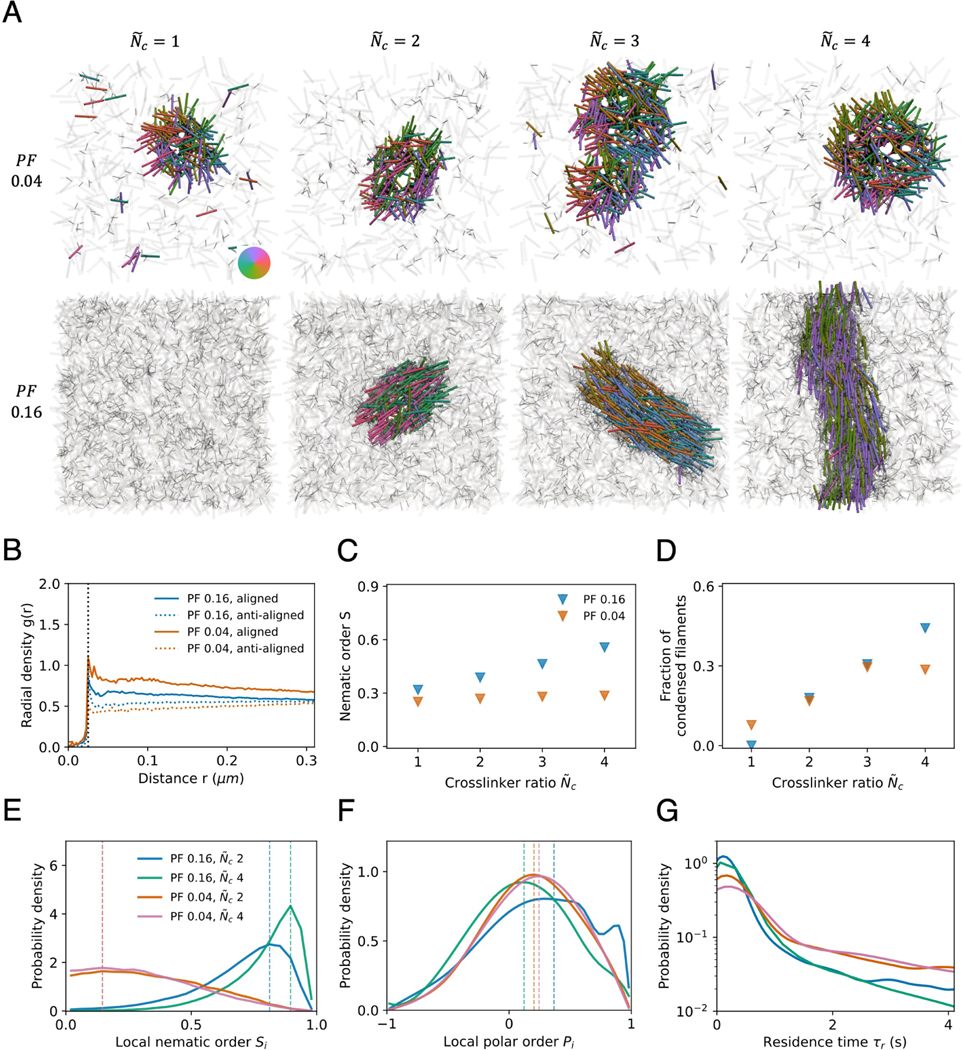
Living systems exhibit self-organization, a phenomenon that enables organisms to perform functions essential for life. The interior of living cells is a crowded environment in which the self-assembly of cytoskeletal networks is spatially constrained by membranes and organelles. Cytoskeletal filaments undergo active condensation in the presence of crosslinking motor proteins. In past studies, confinement has been shown to alter the morphology of active condensates. Here, we perform simulations to explore systems of filaments and crosslinking motors in a variety of confining geometries. We simulate spatial confinement imposed by hard spherical, cylindrical, and planar boundaries. These systems exhibit non-equilibrium condensation behavior where crosslinking motors condense a fraction of the overall filament population, leading to coexistence of vapor and condensed states. We find that the confinement lengthscale modifies the dynamics and condensate morphology. With end-pausing crosslinking motors, filaments self-organize into half asters and fully-symmetric asters under spherical confinement, polarity-sorted bilayers and bottle-brush-like states under cylindrical confinement, and flattened asters under planar confinement. The number of crosslinking motors controls the size and shape of condensates, with flattened asters becoming hollow and ring-like for larger motor number. End pausing plays a key role affecting condensate morphology: systems with end-pausing motors evolve into aster-like condensates while those with non-end-pausing crosslinking motor proteins evolve into disordered clusters and polarity-sorted bundles.
Keywords: active matter; condensation; confinement; crosslinking motors; filaments; microtubules; self-organization.
Conflict of interest statement
CONFLICT OF INTEREST STATEMENT The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Spherical network contraction forms microtubule asters in confinement.Soft Matter. 2018 Feb 14;14(6):901-909. doi: 10.1039/c7sm01718a. Epub 2018 Jan 24. Soft Matter. 2018. PMID: 29364311
-
Macromolecular crowding: chemistry and physics meet biology (Ascona, Switzerland, 10-14 June 2012).Phys Biol. 2013 Aug;10(4):040301. doi: 10.1088/1478-3975/10/4/040301. Epub 2013 Aug 2. Phys Biol. 2013. PMID: 23912807
-
Interplay of self-organization of microtubule asters and crosslinking protein condensates.PNAS Nexus. 2023 Jul 13;2(7):pgad231. doi: 10.1093/pnasnexus/pgad231. eCollection 2023 Jul. PNAS Nexus. 2023. PMID: 37497046 Free PMC article.
-
Active gels: where polymer physics meets cytoskeletal dynamics.Philos Trans A Math Phys Eng Sci. 2006 Dec 15;364(1849):3335-55. doi: 10.1098/rsta.2006.1897. Philos Trans A Math Phys Eng Sci. 2006. PMID: 17090463 Review.
-
Vesicle transport: the role of actin filaments and myosin motors.Microsc Res Tech. 1999 Oct 15;47(2):93-106. doi: 10.1002/(SICI)1097-0029(19991015)47:2<93::AID-JEMT2>3.0.CO;2-P. Microsc Res Tech. 1999. PMID: 10523788 Review.
Cited by
-
Kinesin-5/Cut7 C-terminal tail phosphorylation is essential for microtubule sliding force and bipolar mitotic spindle assembly.Curr Biol. 2024 Oct 21;34(20):4781-4793.e6. doi: 10.1016/j.cub.2024.08.035. Epub 2024 Oct 15. Curr Biol. 2024. PMID: 39413787
References
-
- Gompper G, Winkler RG, Speck T, Solon A, Nardini C, Peruani F, et al. The 2020 motile active matter roadmap. Journal of Physics: Condensed Matter 32 (2020) 193001. - PubMed
-
- Moore JM, Thompson TN, Glaser MA, Betterton MD. Collective motion of driven semiflexible filaments tuned by soft repulsion and stiffness. Soft Matter 16 (2020) 9436–9442. - PubMed
-
- Bricard A, Caussin JB, Desreumaux N, Dauchot O, Bartolo D. Emergence of macroscopic directed motion in populations of motile colloids. Nature 503 (2013) 95–98. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous