

Plant-derived Durvalumab variants show efficient PD-1/PD-L1 blockade and therapeutically favourable FcR binding
- PMID: 38050338
- PMCID: PMC11022803
- DOI: 10.1111/pbi.14260
Plant-derived Durvalumab variants show efficient PD-1/PD-L1 blockade and therapeutically favourable FcR binding
Abstract
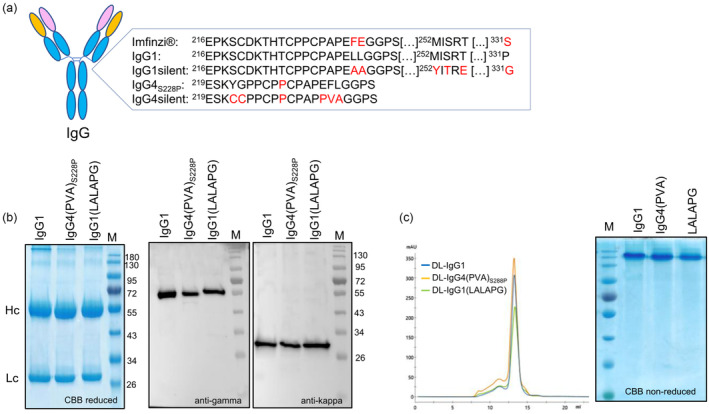
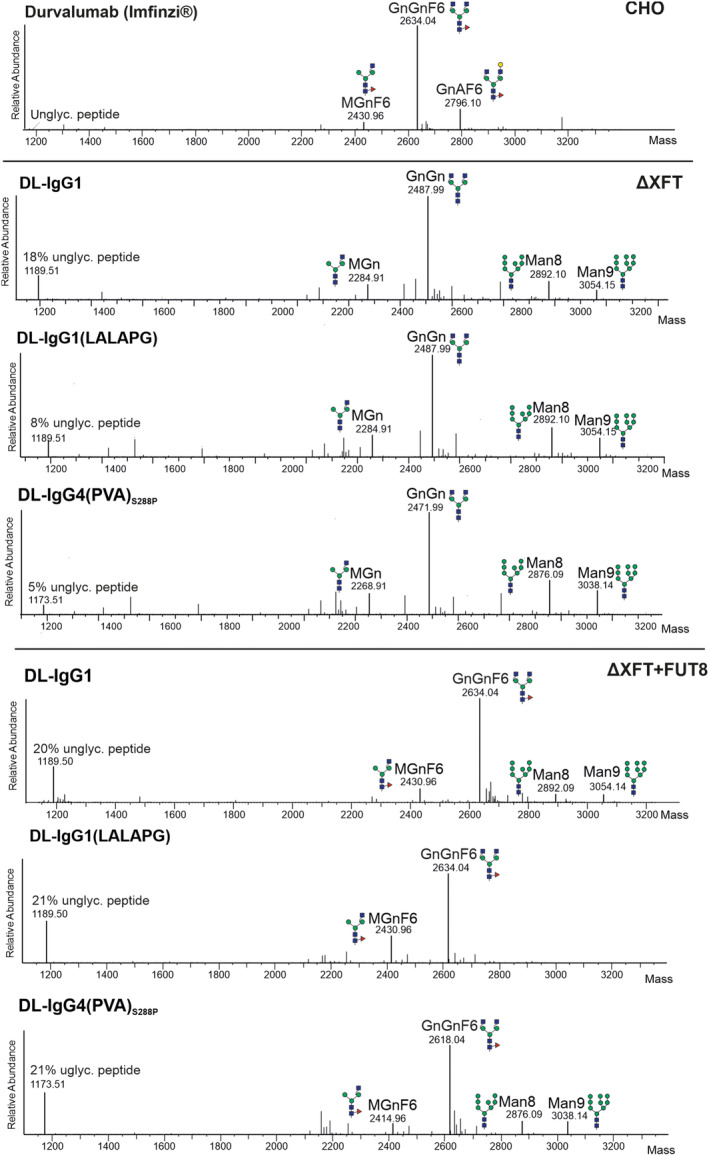
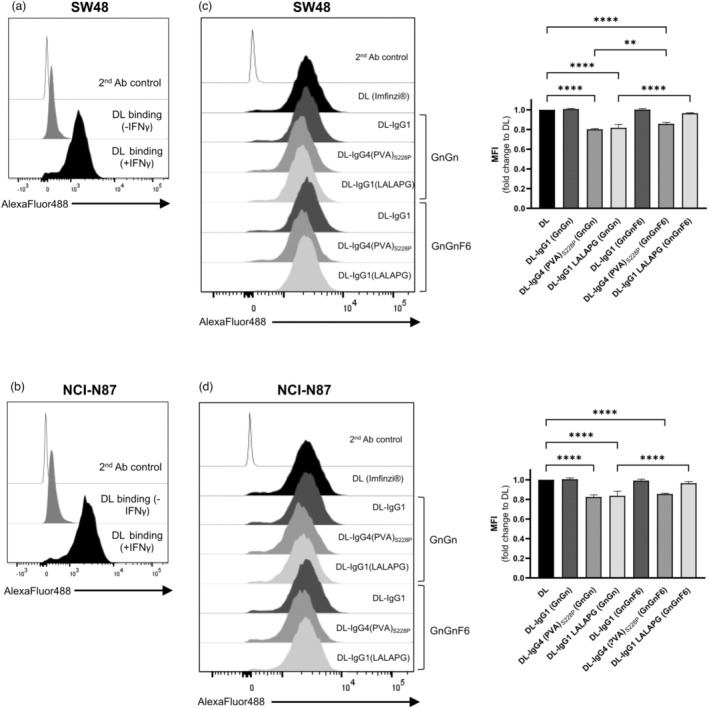
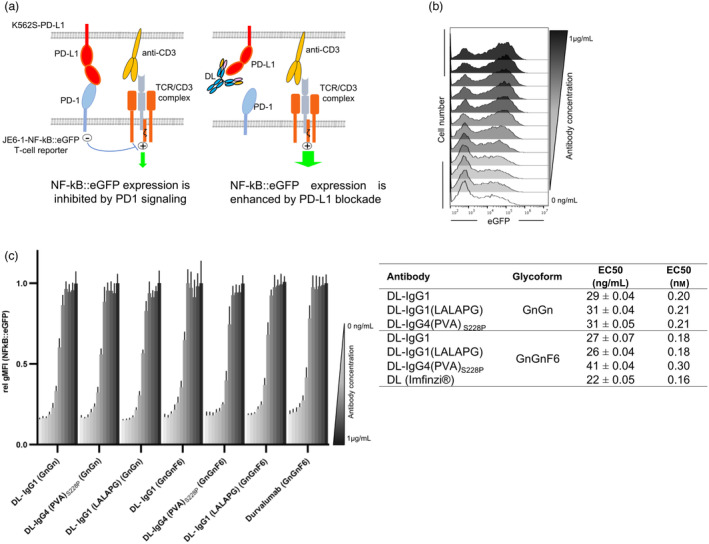
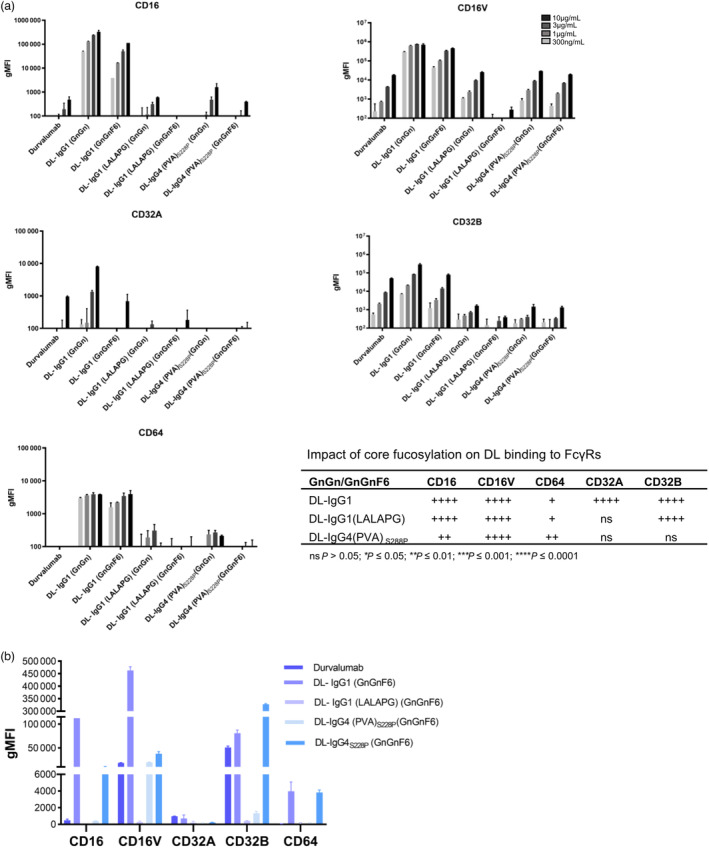
Immune checkpoint blocking therapy targeting the PD-1/PD-L1 inhibitory signalling pathway has produced encouraging results in the treatment of a variety of cancers. Durvalumab (Imfinzi®) targeting PD-L1 is currently used for immunotherapy of several tumour malignancies. The Fc region of this IgG1 antibody has been engineered to reduce FcγR interactions with the aim of enhancing blockade of PD-1/PD-L1 interactions without the depletion of PD-L1-expressing immune cells. Here, we used Nicotiana benthamiana to produce four variants of Durvalumab (DL): wild-type IgG1 and its 'Fc-effector-silent' variant (LALAPG) carrying further modifications to increase antibody half-life (YTE); IgG4S228P and its variant (PVA) with Fc mutations to decrease binding to FcγRI. In addition, DL variants were produced with two distinct glycosylation profiles: afucosylated and decorated with α1,6-core fucose. Plant-derived DL variants were compared to the therapeutic antibody regarding their ability to (i) bind to PD-L1, (ii) block PD-1/PD-L1 inhibitory signalling and (iii) engage with the neonatal Fc receptor (FcRn) and various Fcγ receptors. It was found that plant-derived DL variants bind to recombinant PD-L1 and to PD-L1 expressed in gastrointestinal cancer cells and are able to effectively block its interaction with PD-1 on T cells, thereby enhancing their activation. Furthermore, we show a positive impact of Fc amino acid mutations and core fucosylation on DL's therapeutic potential. Compared to Imfinzi®, DL-IgG1 (LALAPG) and DL-IgG4 (PVA)S228P show lower affinity to CD32B inhibitory receptor which can be therapeutically favourable. Importantly, DL-IgG1 (LALAPG) also shows enhanced binding to FcRn, a key determinant of serum half-life of IgGs.
Keywords: Nicotiana benthamiana; Durvalumab; FcRn; FcγR; PD‐1/PD‐L1; checkpoint inhibitors.
© 2023 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Fc glycoengineering of a PD-L1 antibody harnesses Fcγ receptors for increased antitumor efficacy.Sci Immunol. 2023 Mar 10;8(81):eadd8005. doi: 10.1126/sciimmunol.add8005. Epub 2023 Mar 3. Sci Immunol. 2023. PMID: 36867679
-
Efficient PD-L1 imaging of murine glioblastoma with FUS-aided immunoPET by leveraging FcRn-antibody interaction.Theranostics. 2023 Oct 16;13(15):5584-5596. doi: 10.7150/thno.87168. eCollection 2023. Theranostics. 2023. PMID: 37908736 Free PMC article.
-
Blockade of LAG-3 in PD-L1-Deficient Mice Enhances Clearance of Blood Stage Malaria Independent of Humoral Responses.Front Immunol. 2021 Jan 14;11:576743. doi: 10.3389/fimmu.2020.576743. eCollection 2020. Front Immunol. 2021. PMID: 33519801 Free PMC article.
-
The Next Immune-Checkpoint Inhibitors: PD-1/PD-L1 Blockade in Melanoma.Clin Ther. 2015 Apr 1;37(4):764-82. doi: 10.1016/j.clinthera.2015.02.018. Epub 2015 Mar 29. Clin Ther. 2015. PMID: 25823918 Free PMC article. Review.
-
Durvalumab: an investigational anti-PD-L1 monoclonal antibody for the treatment of urothelial carcinoma.Drug Des Devel Ther. 2018 Jan 23;12:209-215. doi: 10.2147/DDDT.S141491. eCollection 2018. Drug Des Devel Ther. 2018. PMID: 29416316 Free PMC article. Review.
References
-
- Arnold, J.N. , Wormald, M.R. , Sim, R.B. , Rudd, P.M. and Dwek, R.A. (2007) The impact of glycosylation on the biological function and structure of human immunoglobulins. Annu. Rev. Immunol. 25, 21–50. - PubMed
-
- Baumeister, S.H. , Freeman, G.J. , Dranoff, G. and Sharpe, A.H. (2016) Coinhibitory pathways in immunotherapy for cancer. Annu. Rev. Immunol. 34, 539–573. - PubMed
-
- Bellis, S.L. , Reis, C.A. , Varki, A. , Kannagi, R. and Stanley, P. (2022) Glycosylation changes in cancer. In Essentials of Glycobiology ( Varki, A. , Cummings, R.D. , Esko, J.D. , Stanley, P. , Hart, G.W. , Aebi, M. , Mohnen, D. et al., eds), pp. 631–644. Harbor (NY): Cold Spring.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
