

A guide to ERK dynamics, part 2: downstream decoding
- PMID: 38038975
- PMCID: PMC10754290
- DOI: 10.1042/BCJ20230277
A guide to ERK dynamics, part 2: downstream decoding
Abstract
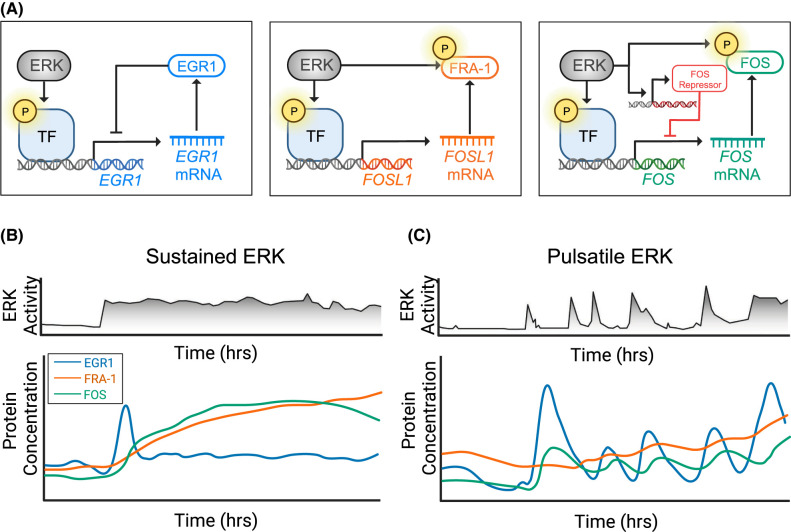
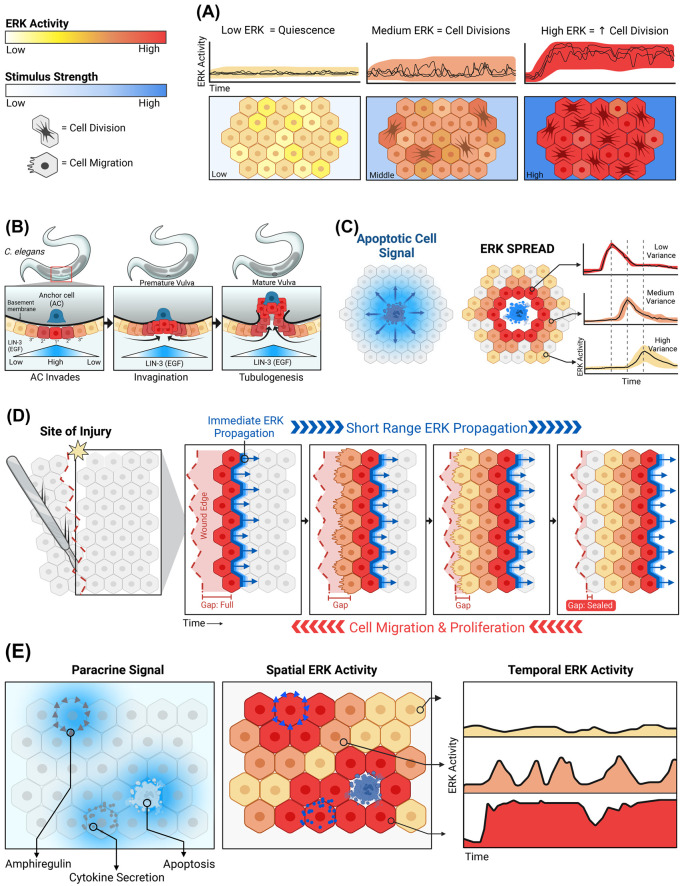
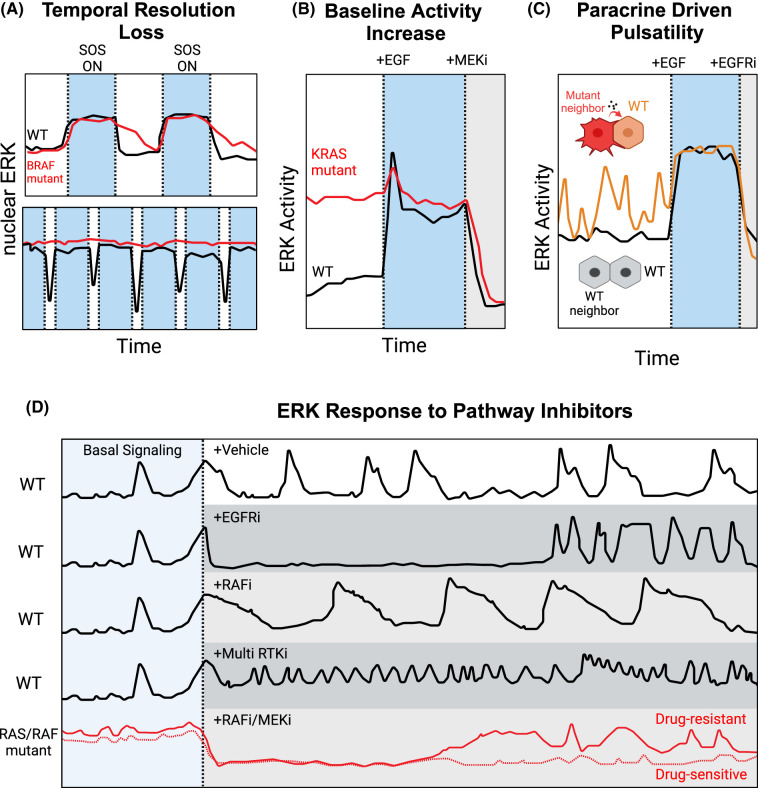
Signaling by the extracellular signal-regulated kinase (ERK) pathway controls many cellular processes, including cell division, death, and differentiation. In this second installment of a two-part review, we address the question of how the ERK pathway exerts distinct and context-specific effects on multiple processes. We discuss how the dynamics of ERK activity induce selective changes in gene expression programs, with insights from both experiments and computational models. With a focus on single-cell biosensor-based studies, we summarize four major functional modes for ERK signaling in tissues: adjusting the size of cell populations, gradient-based patterning, wave propagation of morphological changes, and diversification of cellular gene expression states. These modes of operation are disrupted in cancer and other related diseases and represent potential targets for therapeutic intervention. By understanding the dynamic mechanisms involved in ERK signaling, there is potential for pharmacological strategies that not only simply inhibit ERK, but also restore functional activity patterns and improve disease outcomes.
Keywords: cell proliferation; eukaryotic gene expression; extracellular signal-regulated kinases; gene regulatory networks; receptor tyrosine kinases.
© 2023 The Author(s).
Conflict of interest statement
The authors declare that there are no competing interests associated with the manuscript.
Figures
Similar articles
-
A guide to ERK dynamics, part 1: mechanisms and models.Biochem J. 2023 Dec 13;480(23):1887-1907. doi: 10.1042/BCJ20230276. Biochem J. 2023. PMID: 38038974 Free PMC article.
-
On the inference of ERK signaling dynamics from protein biosensor measurements.Mol Biol Cell. 2023 May 15;34(6):ar60. doi: 10.1091/mbc.E22-10-0476. Epub 2023 Mar 8. Mol Biol Cell. 2023. PMID: 36884295 Free PMC article.
-
Quantification of the Dynamic Phosphorylation Process of ERK Using Stable Isotope Dilution Selective Reaction Monitoring Mass Spectrometry.Proteomics. 2019 Sep;19(17):e1900086. doi: 10.1002/pmic.201900086. Epub 2019 Aug 7. Proteomics. 2019. PMID: 31318149
-
MicroRNAs as regulators of ERK/MAPK pathway: A comprehensive review.Biomed Pharmacother. 2020 Dec;132:110853. doi: 10.1016/j.biopha.2020.110853. Epub 2020 Oct 14. Biomed Pharmacother. 2020. PMID: 33068932 Review.
-
Growth Inhibitory Signaling of the Raf/MEK/ERK Pathway.Int J Mol Sci. 2020 Jul 30;21(15):5436. doi: 10.3390/ijms21155436. Int J Mol Sci. 2020. PMID: 32751750 Free PMC article. Review.
Cited by
-
Asparagine614 Determines the Transport and Function of the Murine Anti-Aging Protein Klotho.Cells. 2024 Oct 21;13(20):1743. doi: 10.3390/cells13201743. Cells. 2024. PMID: 39451260 Free PMC article.
-
Rebound growth of BRAF mutant pediatric glioma cells after MAPKi withdrawal is associated with MAPK reactivation and secretion of microglia-recruiting cytokines.J Neurooncol. 2024 Jun;168(2):317-332. doi: 10.1007/s11060-024-04672-9. Epub 2024 Apr 17. J Neurooncol. 2024. PMID: 38630384 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous