

This is a preprint.
c-Jun Signaling During Initial HSV-1 Infection Modulates Latency to Enhance Later Reactivation in addition to Directly Promoting the Progression to Full Reactivation
- PMID: 37986840
- PMCID: PMC10659354
- DOI: 10.1101/2023.11.10.566462
c-Jun Signaling During Initial HSV-1 Infection Modulates Latency to Enhance Later Reactivation in addition to Directly Promoting the Progression to Full Reactivation
Update in
-
c-Jun signaling during initial HSV-1 infection modulates latency to enhance later reactivation in addition to directly promoting the progression to full reactivation.J Virol. 2024 Feb 20;98(2):e0176423. doi: 10.1128/jvi.01764-23. Epub 2024 Jan 9. J Virol. 2024. PMID: 38193709 Free PMC article.
Abstract
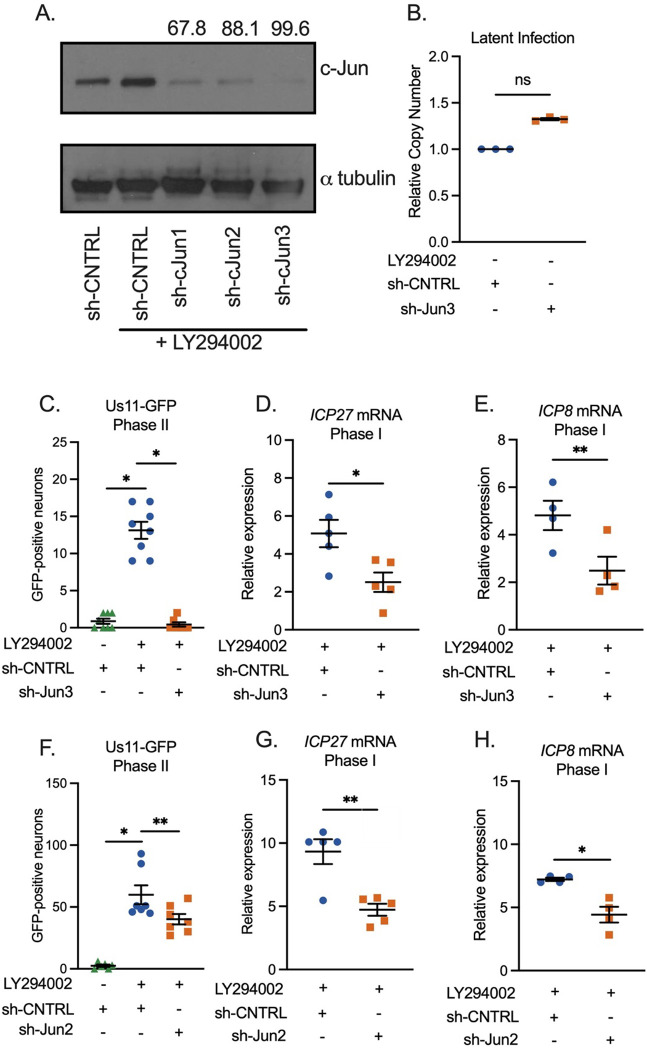
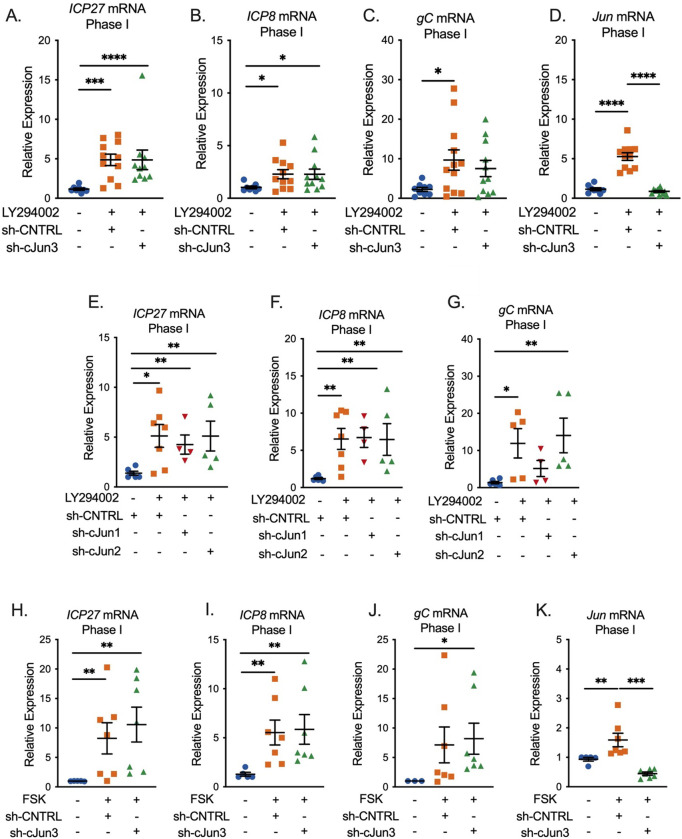
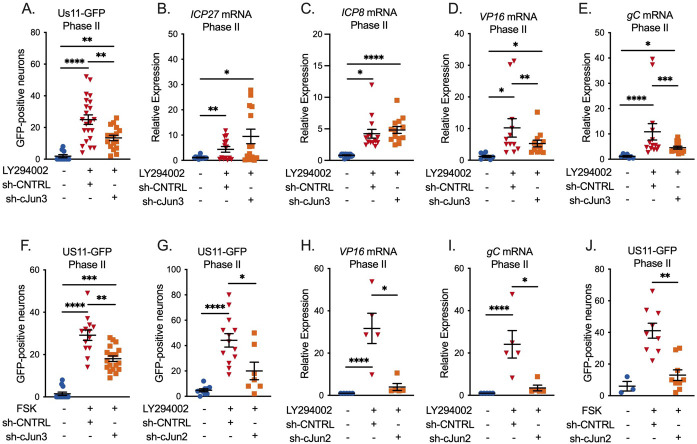
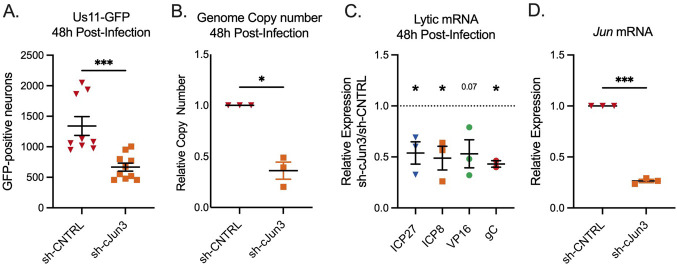
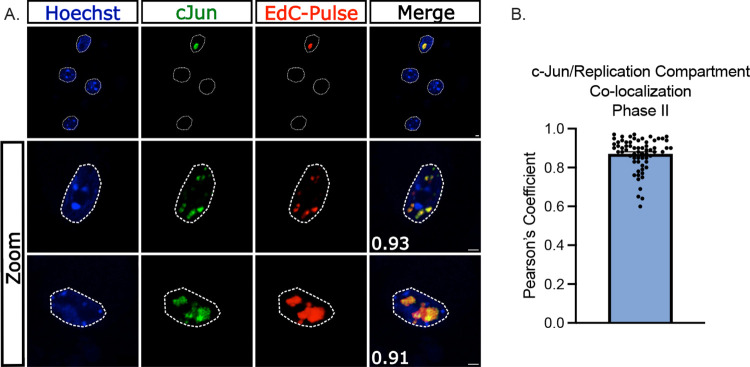
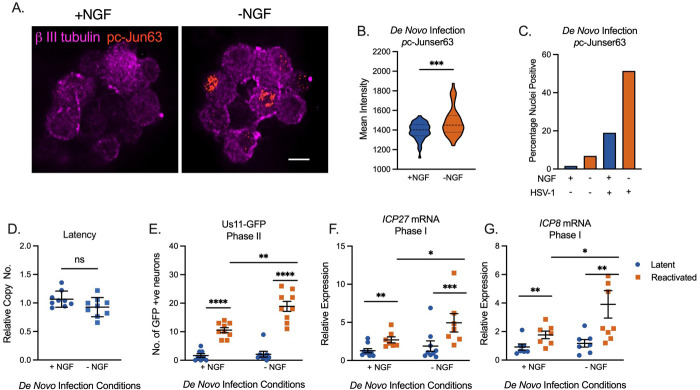
Herpes simplex virus-1 (HSV-1) establishes a latent infection in peripheral neurons and can periodically reactivate to permit transmission and clinical manifestations. Viral transactivators required for lytic infection are largely absent during latent infection and therefore HSV-1 relies on the co-option of neuronal host signaling pathways to initiate its gene expression. Activation of the neuronal c-Jun N-terminal kinase (JNK) cell stress pathway is central to initiating biphasic reactivation in response to multiple stimuli. However, how host factors work with JNK to stimulate the initial wave of gene expression (known as Phase I) or the progression to full, Phase II reactivation remains unclear. Here, we found that c-Jun, the primary target downstream of neuronal JNK cell stress signaling, functions during reactivation but not during the JNK-mediated initiation of Phase I gene expression. Instead, c-Jun was required for the transition from Phase I to full HSV-1 reactivation and was detected in viral replication compartments of reactivating neurons. Interestingly, we also identified a role for both c-Jun and enhanced neuronal stress during initial neuronal infection in promoting a more reactivation-competent form of HSV-1 latency. Therefore, c-Jun functions at multiple stages during HSV latent infection of neurons to promote reactivation. Importantly, by demonstrating that initial infection conditions can contribute to later reactivation abilities, this study highlights the potential for latently infected neurons to maintain a molecular scar of previous exposure to neuronal stressors.
Figures
Similar articles
-
c-Jun signaling during initial HSV-1 infection modulates latency to enhance later reactivation in addition to directly promoting the progression to full reactivation.J Virol. 2024 Feb 20;98(2):e0176423. doi: 10.1128/jvi.01764-23. Epub 2024 Jan 9. J Virol. 2024. PMID: 38193709 Free PMC article.
-
DLK-Dependent Biphasic Reactivation of Herpes Simplex Virus Latency Established in the Absence of Antivirals.J Virol. 2022 Jun 22;96(12):e0050822. doi: 10.1128/jvi.00508-22. Epub 2022 May 24. J Virol. 2022. PMID: 35608347 Free PMC article.
-
Ex Vivo Herpes Simplex Virus Reactivation Involves a Dual Leucine Zipper Kinase-Dependent Wave of Lytic Gene Expression That Is Independent of Histone Demethylase Activity and Viral Genome Synthesis.J Virol. 2022 Jun 22;96(12):e0047522. doi: 10.1128/jvi.00475-22. Epub 2022 May 23. J Virol. 2022. PMID: 35604215 Free PMC article.
-
A comparison of herpes simplex virus type 1 and varicella-zoster virus latency and reactivation.J Gen Virol. 2015 Jul;96(Pt 7):1581-602. doi: 10.1099/vir.0.000128. Epub 2015 Mar 20. J Gen Virol. 2015. PMID: 25794504 Free PMC article. Review.
-
Restarting Lytic Gene Transcription at the Onset of Herpes Simplex Virus Reactivation.J Virol. 2017 Jan 3;91(2):e01419-16. doi: 10.1128/JVI.01419-16. Print 2017 Jan 15. J Virol. 2017. PMID: 27807236 Free PMC article. Review.
References
-
- Knipe DM, Cliffe A. 2008. Chromatin control of herpes simplex virus lytic and latent infection. Nat Rev Microbiol 6:211–21. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous