

FHL1 promotes chikungunya and o'nyong-nyong virus infection and pathogenesis with implications for alphavirus vaccine design
- PMID: 37884534
- PMCID: PMC10603155
- DOI: 10.1038/s41467-023-42330-2
FHL1 promotes chikungunya and o'nyong-nyong virus infection and pathogenesis with implications for alphavirus vaccine design
Abstract
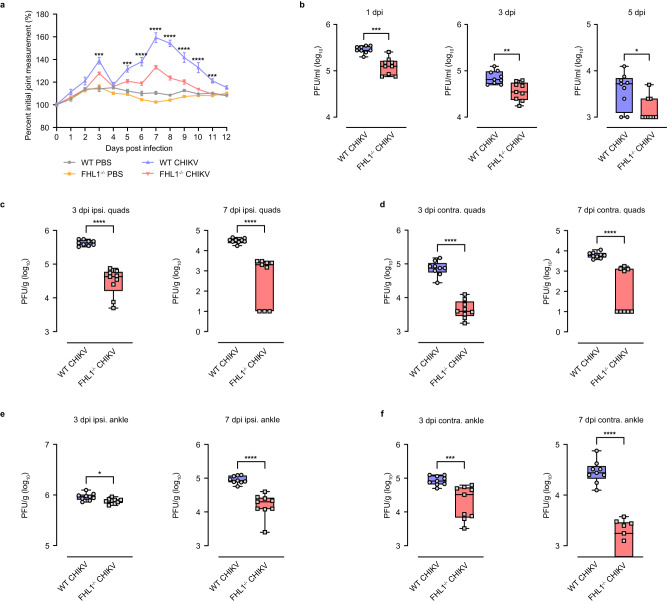
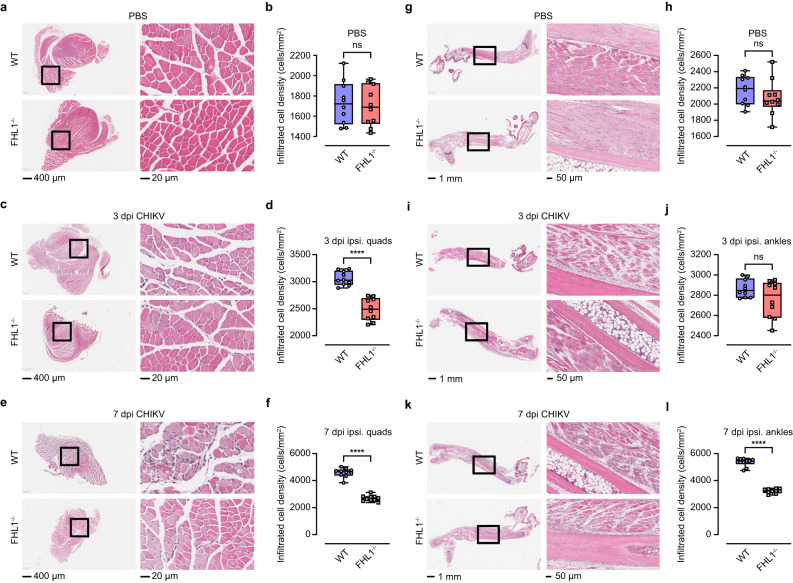
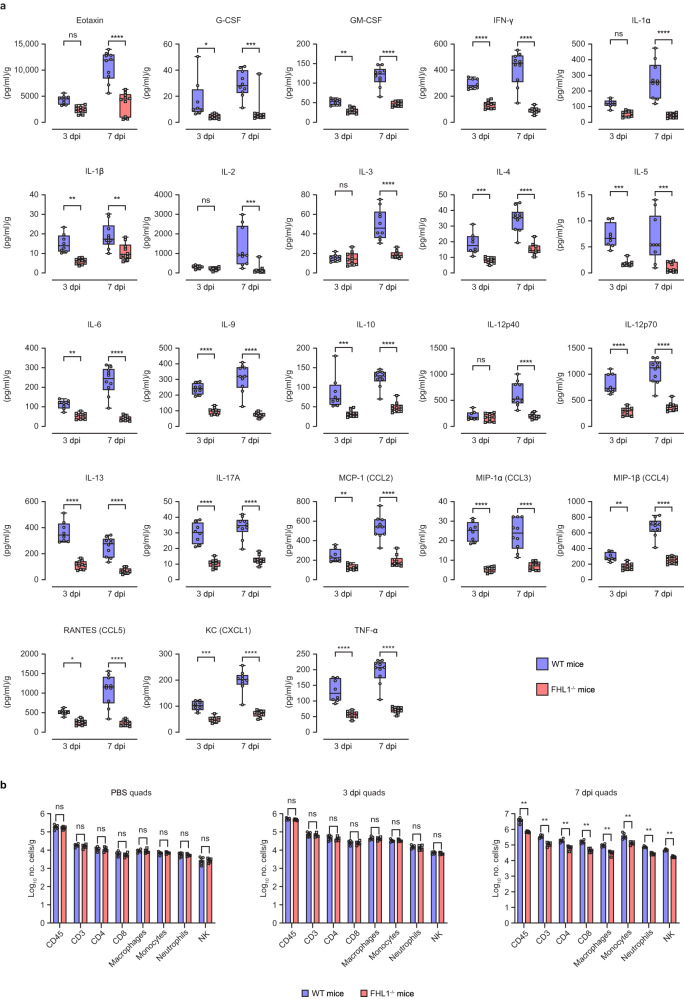
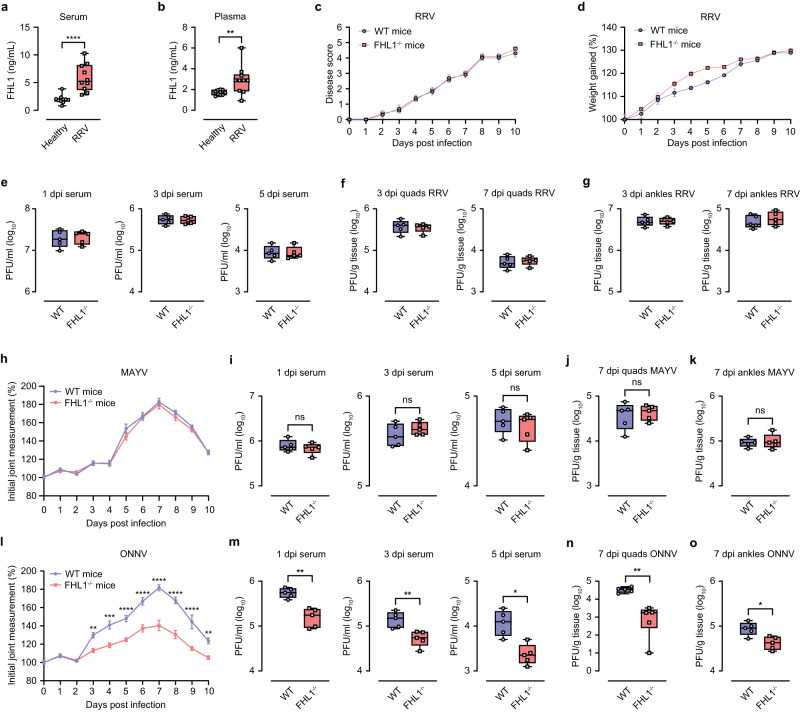
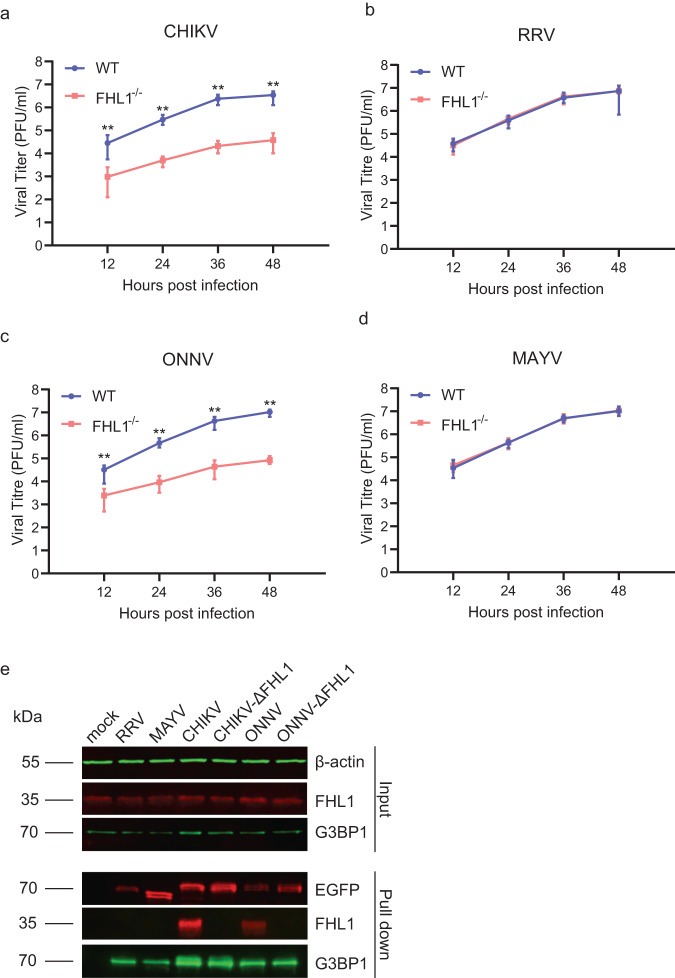
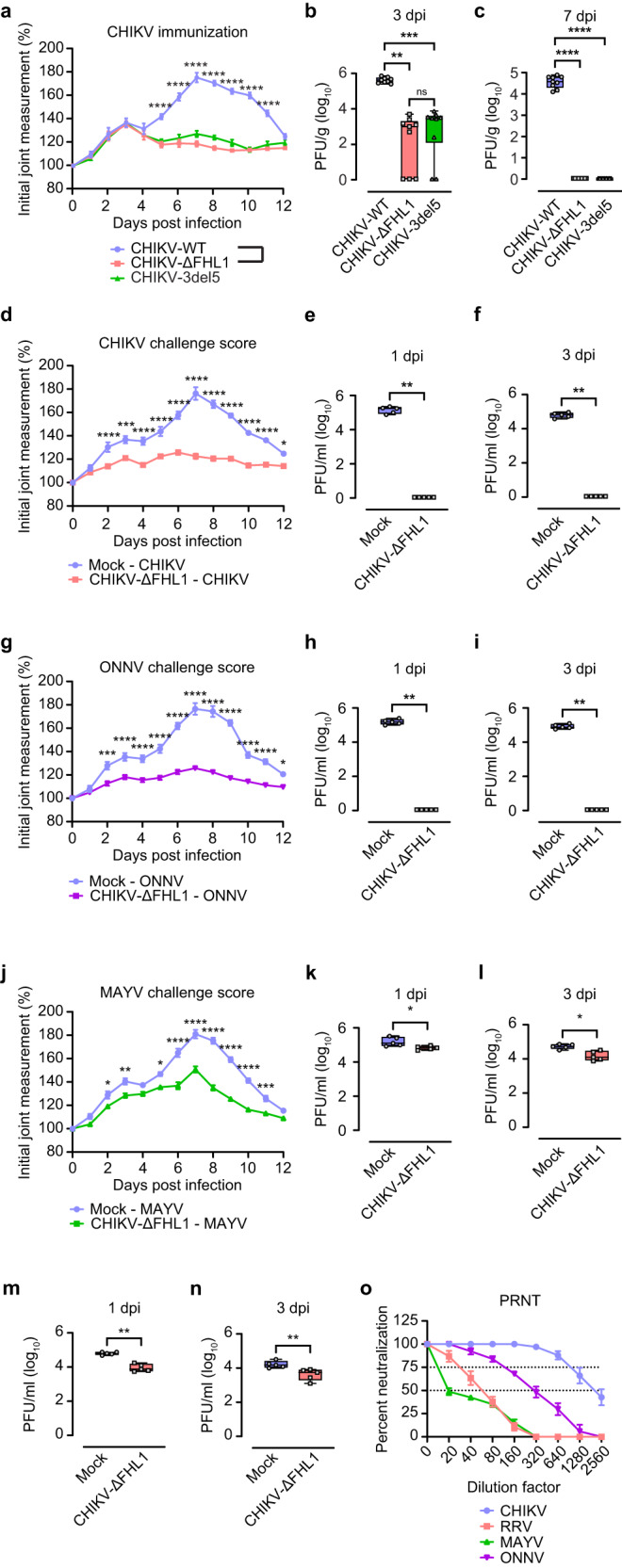
Arthritogenic alphaviruses are positive-strand RNA viruses that cause debilitating musculoskeletal diseases affecting millions worldwide. A recent discovery identified the four-and-a-half-LIM domain protein 1 splice variant A (FHL1A) as a crucial host factor interacting with the hypervariable domain (HVD) of chikungunya virus (CHIKV) nonstructural protein 3 (nsP3). Here, we show that acute and chronic chikungunya disease in humans correlates with elevated levels of FHL1. We generated FHL1-/- mice, which when infected with CHIKV or o'nyong-nyong virus (ONNV) displayed reduced arthritis and myositis, fewer immune infiltrates, and reduced proinflammatory cytokine/chemokine outputs, compared to infected wild-type (WT) mice. Interestingly, disease signs were comparable in FHL1-/- and WT mice infected with arthritogenic alphaviruses Ross River virus (RRV) or Mayaro virus (MAYV). This aligns with pull-down assay data, which showed the ability of CHIKV and ONNV nsP3 to interact with FHL1, while RRV and MAYV nsP3s did not. We engineered a CHIKV mutant unable to bind FHL1 (CHIKV-ΔFHL1), which was avirulent in vivo. Following inoculation with CHIKV-ΔFHL1, mice were protected from disease upon challenge with CHIKV and ONNV, and viraemia was significantly reduced in RRV- and MAYV-challenged mice. Targeting FHL1-binding as an approach to vaccine design could lead to breakthroughs in mitigating alphaviral disease.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
FHL1 is a major host factor for chikungunya virus infection.Nature. 2019 Oct;574(7777):259-263. doi: 10.1038/s41586-019-1578-4. Epub 2019 Sep 25. Nature. 2019. PMID: 31554973
-
Structural and Functional Characterization of Host FHL1 Protein Interaction with Hypervariable Domain of Chikungunya Virus nsP3 Protein.J Virol. 2020 Dec 9;95(1):e01672-20. doi: 10.1128/JVI.01672-20. Print 2020 Dec 9. J Virol. 2020. PMID: 33055253 Free PMC article.
-
Role of MXRA8 in Ross River Virus Disease Pathogenesis.mBio. 2023 Apr 25;14(2):e0058823. doi: 10.1128/mbio.00588-23. Epub 2023 Apr 10. mBio. 2023. PMID: 37036079 Free PMC article.
-
GloPID-R report on chikungunya, o'nyong-nyong and Mayaro virus, part 2: Epidemiological distribution of o'nyong-nyong virus.Antiviral Res. 2019 Dec;172:104611. doi: 10.1016/j.antiviral.2019.104611. Epub 2019 Sep 20. Antiviral Res. 2019. PMID: 31545982 Review.
-
GloPID-R report on chikungunya, o'nyong-nyong and Mayaro virus, part 5: Entomological aspects.Antiviral Res. 2020 Feb;174:104670. doi: 10.1016/j.antiviral.2019.104670. Epub 2019 Dec 5. Antiviral Res. 2020. PMID: 31812638 Review.
Cited by
-
Alphavirus nsP3 organizes into tubular scaffolds essential for infection and the cytoplasmic granule architecture.Nat Commun. 2024 Sep 16;15(1):8106. doi: 10.1038/s41467-024-51952-z. Nat Commun. 2024. PMID: 39285216 Free PMC article.
-
Emergence of ECSA-IOL E1-K211E/E2-V264A Lineage of Chikungunya virus during Malaysian 2021 outbreak.BMC Infect Dis. 2024 Oct 24;24(1):1199. doi: 10.1186/s12879-024-10102-y. BMC Infect Dis. 2024. PMID: 39448916 Free PMC article.
-
Chronic chikungunya disease (CCD): clinical insights, immunopathogenesis and therapeutic perspectives.QJM. 2024 Jul 1;117(7):489-494. doi: 10.1093/qjmed/hcae028. QJM. 2024. PMID: 38377410 Free PMC article. Review.
-
Pathogenicity and virulence of O'nyong-nyong virus: A less studied Togaviridae with pandemic potential.Virulence. 2024 Dec;15(1):2355201. doi: 10.1080/21505594.2024.2355201. Epub 2024 May 26. Virulence. 2024. PMID: 38797948 Free PMC article. Review.
-
LDL receptor in alphavirus entry: structural analysis and implications for antiviral therapy.Nat Commun. 2024 Jun 8;15(1):4906. doi: 10.1038/s41467-024-49301-1. Nat Commun. 2024. PMID: 38851803 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
