

This is a preprint.
Structural and dynamic changes in P-Rex1 upon activation by PIP3 and inhibition by IP4
- PMID: 37745379
- PMCID: PMC10515948
- DOI: 10.1101/2023.09.15.557836
Structural and dynamic changes in P-Rex1 upon activation by PIP3 and inhibition by IP4
Update in
-
Structural and dynamic changes in P-Rex1 upon activation by PIP3 and inhibition by IP4.Elife. 2024 Jul 31;12:RP92822. doi: 10.7554/eLife.92822. Elife. 2024. PMID: 39082940 Free PMC article.
Abstract
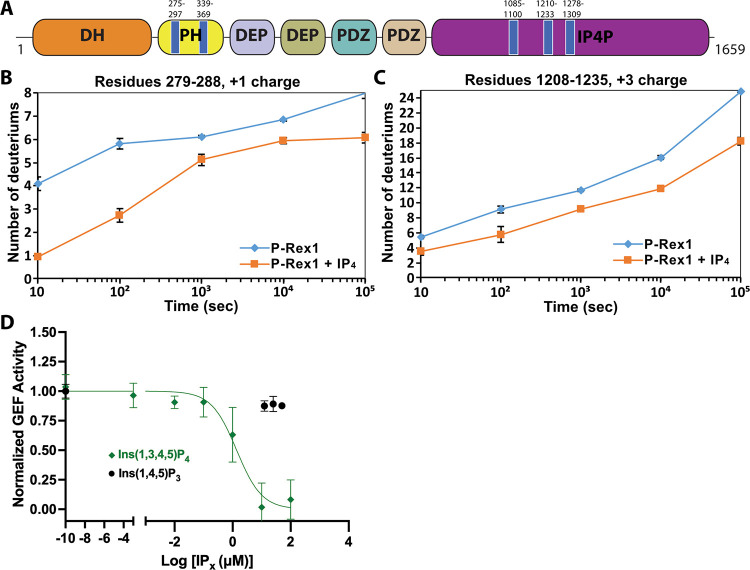
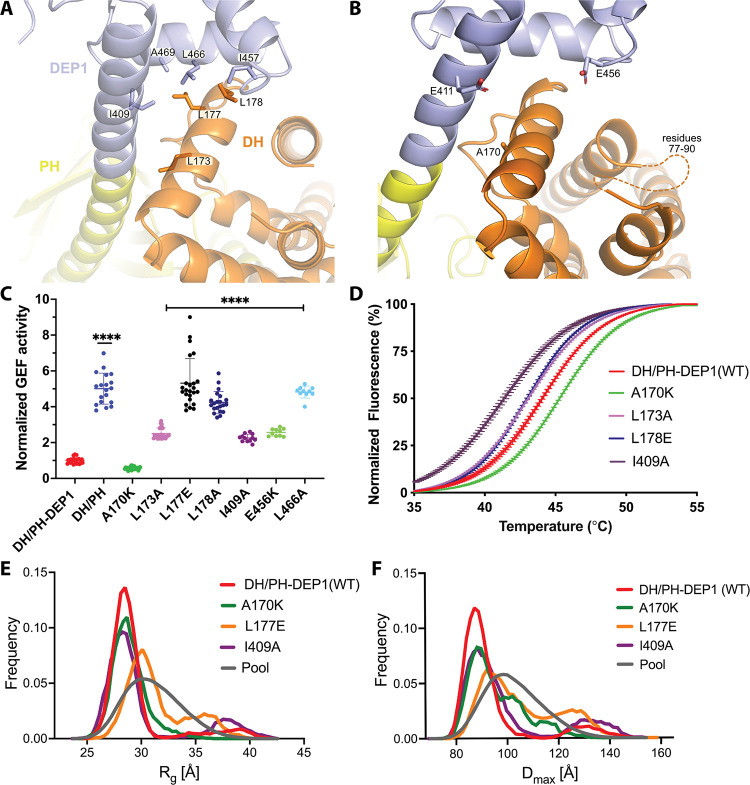
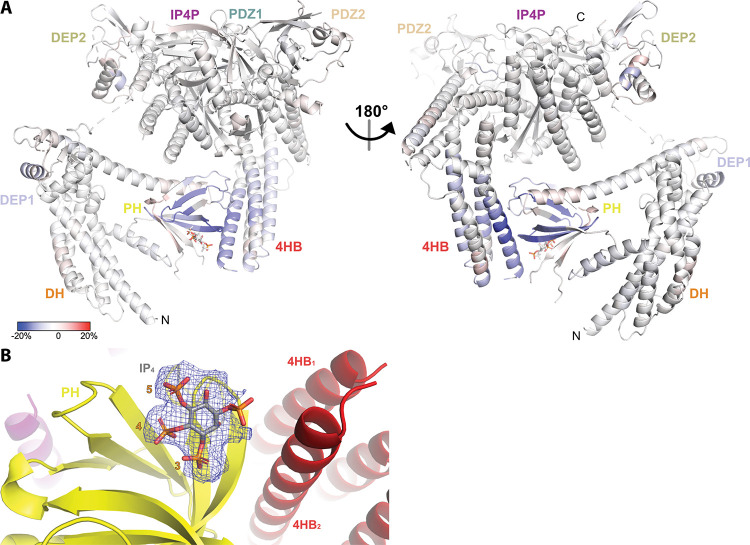
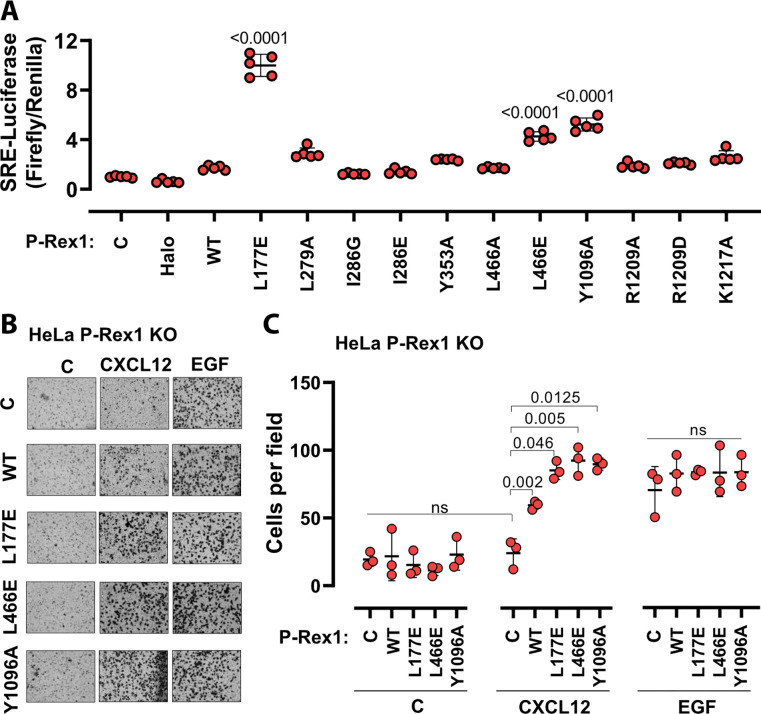
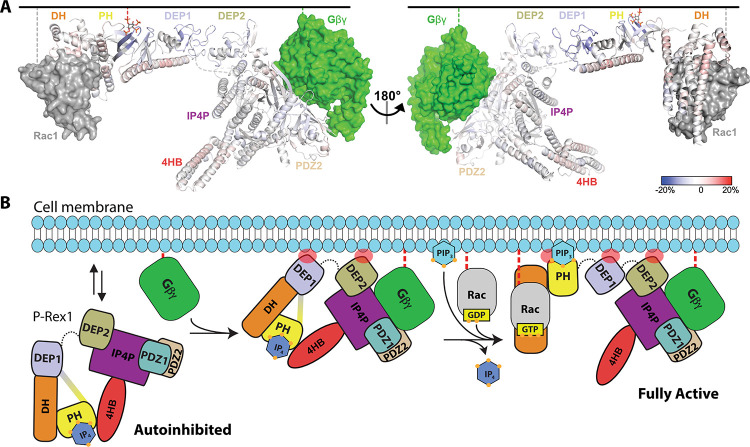
PIP3-dependent Rac exchanger 1 (P-Rex1) is abundantly expressed in neutrophils and plays central roles in chemotaxis and cancer metastasis by serving as a guanine-nucleotide exchange factor (GEF) for Rac. The enzyme is synergistically activated by PIP3 and the heterotrimeric Gβγ subunits, but mechanistic details remain poorly understood. While investigating the regulation of P-Rex1 by PIP3, we discovered that Ins(1,3,4,5)P4 (IP4) inhibits P-Rex1 activity and induces large decreases in backbone dynamics in diverse regions of the protein. Cryo-electron microscopy analysis of the P-Rex1·IP4 complex revealed a conformation wherein the pleckstrin homology (PH) domain occludes the active site of the Dbl homology (DH) domain. This configuration is stabilized by interactions between the first DEP domain (DEP1) and the DH domain and between the PH domain and a 4-helix bundle (4HB) subdomain that extends from the C-terminal domain of P-Rex1. Disruption of the DH-DEP1 interface in a DH/PH-DEP1 fragment enhanced activity and led to a more extended conformation in solution, whereas mutations that constrain the occluded conformation led to decreased GEF activity. Variants of full-length P-Rex1 in which the DH-DEP1 and PH-4HB interfaces were disturbed exhibited enhanced activity during chemokine-induced cell migration, confirming that the observed structure represents the autoinhibited state in living cells. Interactions with PIP3-containing liposomes led to disruption of these interfaces and increased dynamics protein-wide. Our results further suggest that inositol phosphates such as IP4 help to inhibit basal P-Rex1 activity in neutrophils, similar to their inhibitory effects on phosphatidylinositol-3-kinase.
Figures

Similar articles
-
Structural and dynamic changes in P-Rex1 upon activation by PIP3 and inhibition by IP4.Elife. 2024 Jul 31;12:RP92822. doi: 10.7554/eLife.92822. Elife. 2024. PMID: 39082940 Free PMC article.
-
The first DEP domain of the RhoGEF P-Rex1 autoinhibits activity and contributes to membrane binding.J Biol Chem. 2020 Sep 4;295(36):12635-12647. doi: 10.1074/jbc.RA120.014534. Epub 2020 Jul 13. J Biol Chem. 2020. PMID: 32661198 Free PMC article.
-
Discovery of Small Molecules That Target the Phosphatidylinositol (3,4,5) Trisphosphate (PIP3)-Dependent Rac Exchanger 1 (P-Rex1) PIP3-Binding Site and Inhibit P-Rex1-Dependent Functions in Neutrophils.Mol Pharmacol. 2020 Mar;97(3):226-236. doi: 10.1124/mol.119.117556. Epub 2020 Jan 3. Mol Pharmacol. 2020. PMID: 31900312 Free PMC article.
-
The Rho guanine nucleotide exchange factor P-Rex1 as a potential drug target for cancer metastasis and inflammatory diseases.Pharmacol Res. 2020 Mar;153:104676. doi: 10.1016/j.phrs.2020.104676. Epub 2020 Jan 30. Pharmacol Res. 2020. PMID: 32006571 Review.
-
Regulation and function of P-Rex family Rac-GEFs.Small GTPases. 2015;6(2):49-70. doi: 10.4161/21541248.2014.973770. Epub 2015 May 11. Small GTPases. 2015. PMID: 25961466 Free PMC article. Review.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous