

This is a preprint.
Regulatory mechanisms controlling store-operated calcium entry
- PMID: 37744466
- PMCID: PMC10516112
Regulatory mechanisms controlling store-operated calcium entry
Update in
-
Regulatory mechanisms controlling store-operated calcium entry.Front Physiol. 2023 Dec 19;14:1330259. doi: 10.3389/fphys.2023.1330259. eCollection 2023. Front Physiol. 2023. PMID: 38169682 Free PMC article. Review.
Abstract
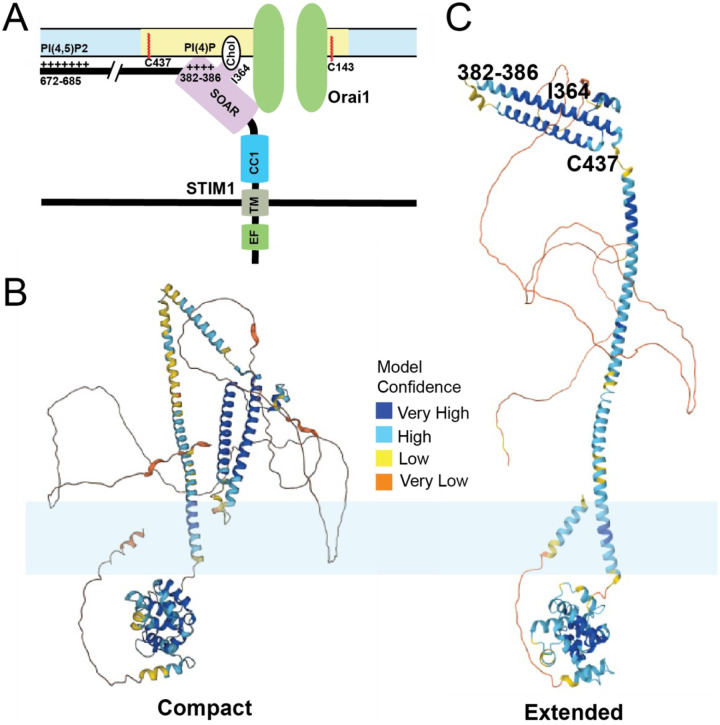
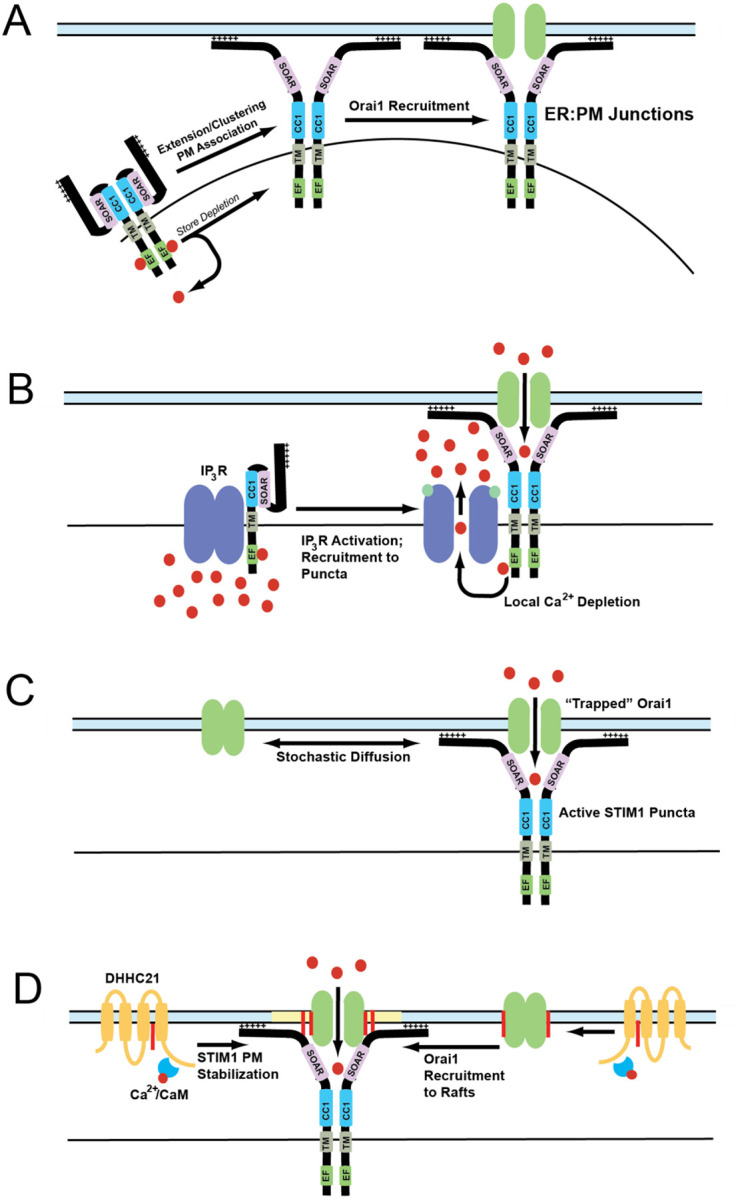
Calcium influx through plasma membrane ion channels is crucial for many events in cellular physiology. Cell surface stimuli lead to the production of inositol 1,4,5-trisphosphate (IP3), which binds to IP3 receptors (IP3R) in the endoplasmic reticulum (ER) to release calcium pools from the ER lumen. This leads to the depletion of ER calcium pools, which has been termed store depletion. Store depletion leads to the dissociation of calcium ions from the EF-hand motif of the ER calcium sensor Stromal Interaction Molecule 1 (STIM1). This leads to a conformational change in STIM1, which helps it to interact with the plasma membrane (PM) at ER:PM junctions. At these ER:PM junctions, STIM1 binds to and activates a calcium channel known as Orai1 to form calcium-release activated calcium (CRAC) channels. Activation of Orai1 leads to calcium influx, known as store-operated calcium entry (SOCE). In addition to Orai1 and STIM1, the homologs of Orai1 and STIM1, such as Orai2/3 and STIM2, also play a crucial role in calcium homeostasis. The influx of calcium through the Orai channel activates a calcium current that has been termed the CRAC current. CRAC channels form multimers and cluster together in large macromolecular assemblies termed "puncta". How CRAC channels form puncta has been contentious since their discovery. In this review, we will outline the history of SOCE, the molecular players involved in this process, as well as the models that have been proposed to explain this critical mechanism in cellular physiology.
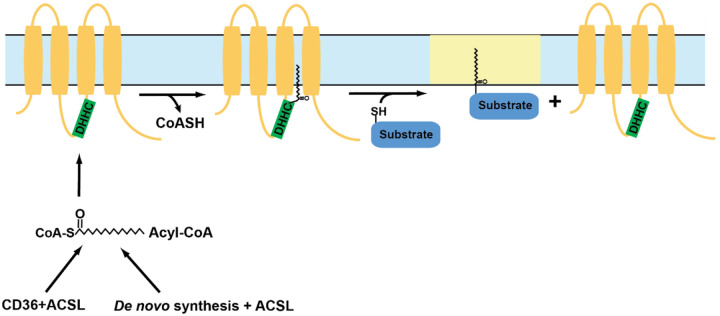
Keywords: calcium; dhhc21; immune diseases; orai1; s-acylation; stim1; store-operated calcium entry.
Conflict of interest statement
Conflict of interest: The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be considered as potential conflict of interest.
Figures

Similar articles
-
Regulatory mechanisms controlling store-operated calcium entry.Front Physiol. 2023 Dec 19;14:1330259. doi: 10.3389/fphys.2023.1330259. eCollection 2023. Front Physiol. 2023. PMID: 38169682 Free PMC article. Review.
-
Store-Independent Orai Channels Regulated by STIM.In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 11. In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 11. PMID: 30299650 Free Books & Documents. Review.
-
Modulation of Orai1 and STIM1 by Cellular Factors.In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 4. In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 4. PMID: 30299655 Free Books & Documents. Review.
-
Studies of Structure-Function and Subunit Composition of Orai/STIM Channel.In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 2. In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 2. PMID: 30299645 Free Books & Documents. Review.
-
Regulation and Role of Store-Operated Ca2+ Entry in Cellular Proliferation.In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 12. In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 12. PMID: 30299656 Free Books & Documents. Review.
References
-
- ALAVIZARGAR A., BERTI C., EJTEHADI M. R. & FURINI S. 2018. Molecular Dynamics Simulations of Orai Reveal How the Third Transmembrane Segment Contributes to Hydration and Ca(2+) Selectivity in Calcium Release-Activated Calcium Channels. J Phys Chem B, 122, 4407–4417. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources