

Structure-based design of a single-chain triple-disulfide-stabilized fusion-glycoprotein trimer that elicits high-titer neutralizing responses against human metapneumovirus
- PMID: 37738240
- PMCID: PMC10516418
- DOI: 10.1371/journal.ppat.1011584
Structure-based design of a single-chain triple-disulfide-stabilized fusion-glycoprotein trimer that elicits high-titer neutralizing responses against human metapneumovirus
Abstract
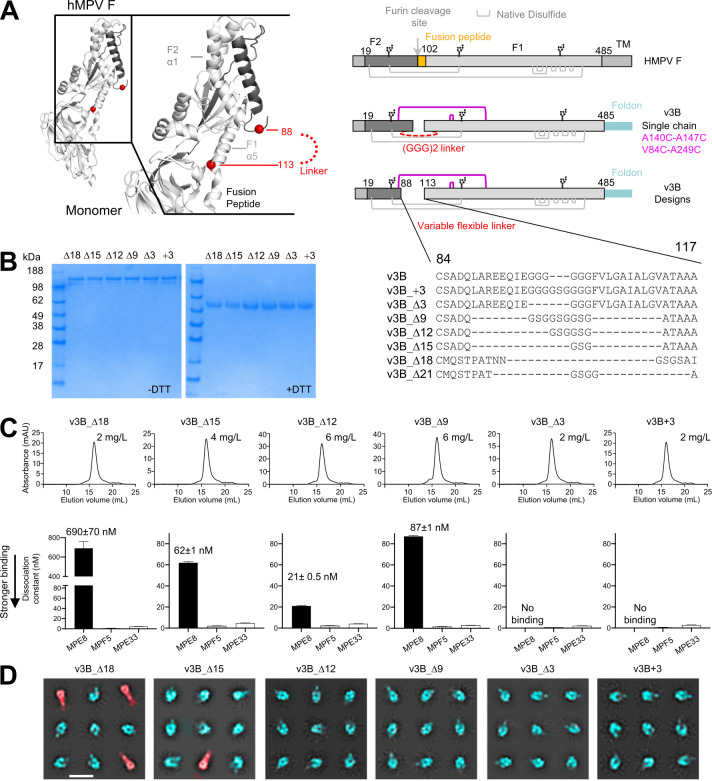
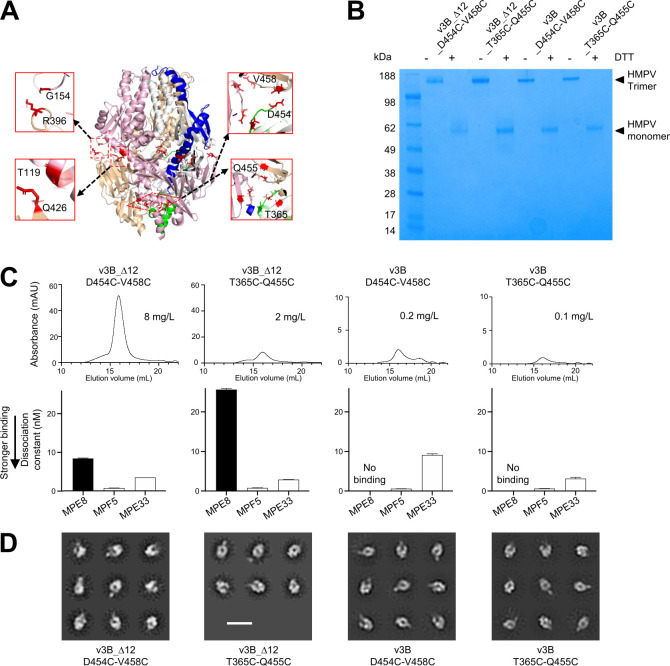
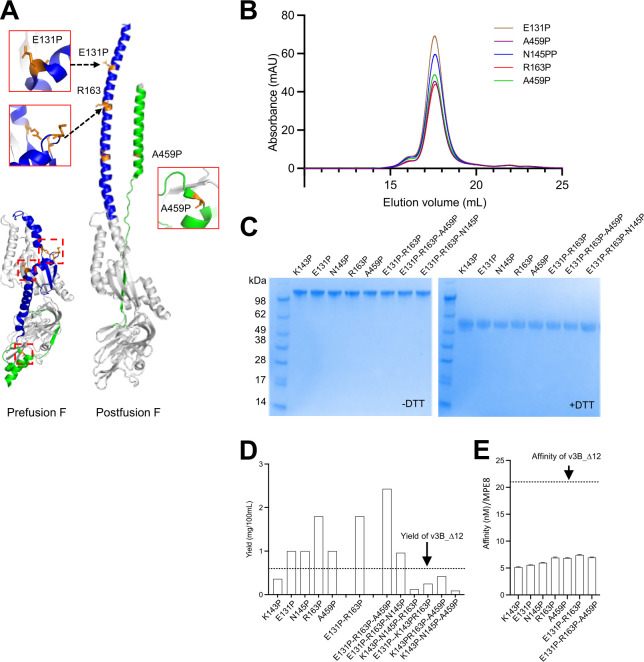
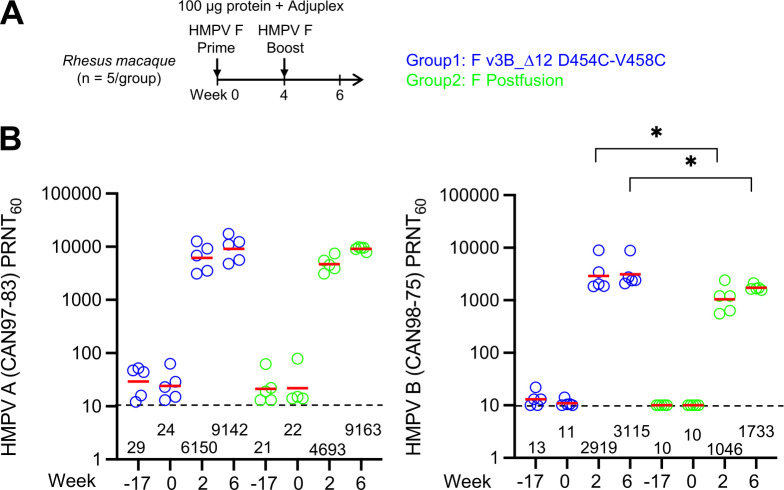
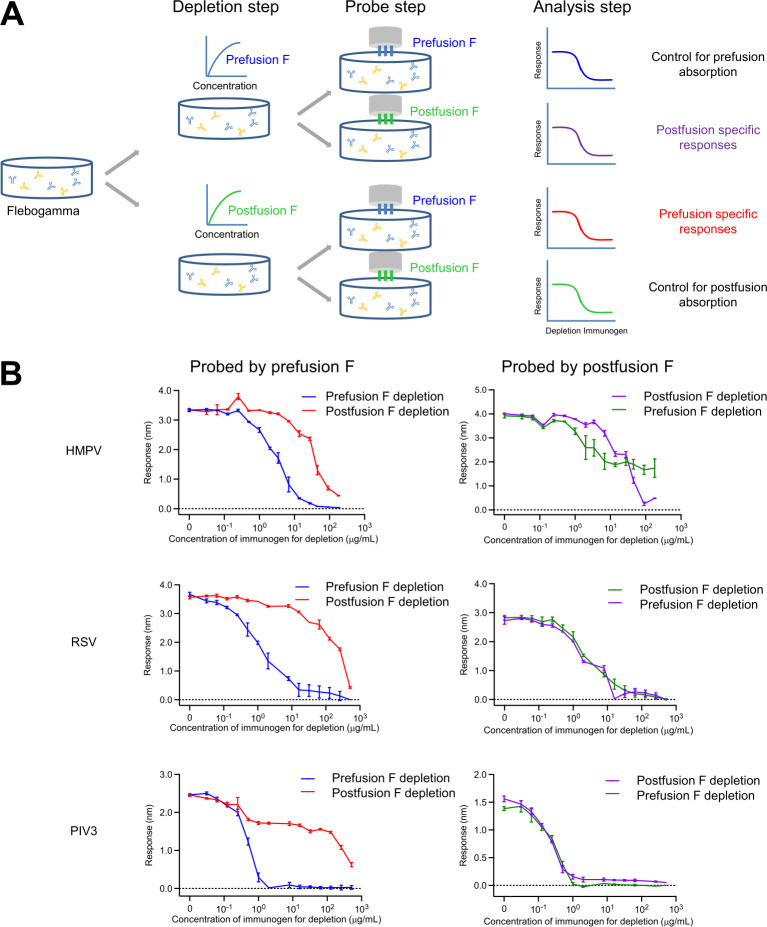
The Pneumoviridae family of viruses includes human metapneumovirus (HMPV) and respiratory syncytial virus (RSV). The closely related Paramyxoviridae family includes parainfluenza viruses (PIVs). These three viral pathogens cause acute respiratory tract infections with substantial disease burden in the young, the elderly, and the immune-compromised. While promising subunit vaccines are being developed with prefusion-stabilized forms of the fusion glycoproteins (Fs) of RSV and PIVs, for which neutralizing titers elicited by the prefusion (pre-F) conformation of F are much higher than for the postfusion (post-F) conformation, with HMPV, pre-F and post-F immunogens described thus far elicit similar neutralizing responses, and it has been unclear which conformation, pre-F or post-F, would be the most effective HMPV F-vaccine immunogen. Here, we investigate the impact of further stabilizing HMPV F in the pre-F state. We replaced the furin-cleavage site with a flexible linker, creating a single chain F that yielded increased amounts of pre-F stabilized trimers, enabling the generation and assessment of F trimers stabilized by multiple disulfide bonds. Introduced prolines could increase both expression yields and antigenic recognition by the pre-F specific antibody, MPE8. The cryo-EM structure of a triple disulfide-stabilized pre-F trimer with the variable region of antibody MPE8 at 3.25-Å resolution confirmed the formation of designed disulfides and provided structural details on the MPE8 interface. Immunogenicity assessments in naïve mice showed the triple disulfide-stabilized pre-F trimer could elicit high titer neutralization, >10-fold higher than elicited by post-F. Immunogenicity assessments in pre-exposed rhesus macaques showed the triple disulfide-stabilized pre-F could recall high neutralizing titers after a single immunization, with little discrimination in the recall response between pre-F and post-F immunogens. However, the triple disulfide-stabilized pre-F adsorbed HMPV-directed responses from commercially available pooled human immunoglobulin more fully than post-F. Collectively, these results suggest single-chain triple disulfide-stabilized pre-F trimers to be promising HMPV-vaccine antigens.
Copyright: This is an open access article, free of all copyright, and may be freely reproduced, distributed, transmitted, modified, built upon, or otherwise used by anyone for any lawful purpose. The work is made available under the Creative Commons CC0 public domain dedication.
Conflict of interest statement
LO, BZ, TZ, WPK, YT, JRM, UJB, and PDK are inventors on a U.S. Patent Application filed on their behalf by the National Institutes of Health. The other authors declare no competing interests.
Figures


Similar articles
-
Rational design of uncleaved prefusion-closed trimer vaccines for human respiratory syncytial virus and metapneumovirus.Nat Commun. 2024 Nov 16;15(1):9939. doi: 10.1038/s41467-024-54287-x. Nat Commun. 2024. PMID: 39550381 Free PMC article.
-
Interprotomer disulfide-stabilized variants of the human metapneumovirus fusion glycoprotein induce high titer-neutralizing responses.Proc Natl Acad Sci U S A. 2021 Sep 28;118(39):e2106196118. doi: 10.1073/pnas.2106196118. Proc Natl Acad Sci U S A. 2021. PMID: 34551978 Free PMC article.
-
Structure-based design of a quadrivalent fusion glycoprotein vaccine for human parainfluenza virus types 1-4.Proc Natl Acad Sci U S A. 2018 Nov 27;115(48):12265-12270. doi: 10.1073/pnas.1811980115. Epub 2018 Nov 12. Proc Natl Acad Sci U S A. 2018. PMID: 30420505 Free PMC article.
-
Engineering, Structure and Immunogenicity of the Human Metapneumovirus F Protein in the Postfusion Conformation.PLoS Pathog. 2016 Sep 9;12(9):e1005859. doi: 10.1371/journal.ppat.1005859. eCollection 2016 Sep. PLoS Pathog. 2016. PMID: 27611367 Free PMC article.
-
Structural basis for respiratory syncytial virus and human metapneumovirus neutralization.Curr Opin Virol. 2023 Aug;61:101337. doi: 10.1016/j.coviro.2023.101337. Curr Opin Virol. 2023. PMID: 37544710 Free PMC article. Review.
Cited by
-
Engineered dityrosine-bonding of the RSV prefusion F protein imparts stability and potency advantages.Nat Commun. 2024 Mar 14;15(1):2202. doi: 10.1038/s41467-024-46295-8. Nat Commun. 2024. PMID: 38485927 Free PMC article.
-
Rational design of uncleaved prefusion-closed trimer vaccines for human respiratory syncytial virus and metapneumovirus.Nat Commun. 2024 Nov 16;15(1):9939. doi: 10.1038/s41467-024-54287-x. Nat Commun. 2024. PMID: 39550381 Free PMC article.
-
Efficacious human metapneumovirus vaccine based on AI-guided engineering of a closed prefusion trimer.Nat Commun. 2024 Jul 25;15(1):6270. doi: 10.1038/s41467-024-50659-5. Nat Commun. 2024. PMID: 39054318 Free PMC article.
-
Structural characterization of M8C10, a neutralizing antibody targeting a highly conserved prefusion-specific epitope on the metapneumovirus fusion trimerization interface.J Virol. 2023 Dec 21;97(12):e0105223. doi: 10.1128/jvi.01052-23. Epub 2023 Nov 30. J Virol. 2023. PMID: 38032197 Free PMC article.
-
A tale of two fusion proteins: understanding the metastability of human respiratory syncytial virus and metapneumovirus and implications for rational design of uncleaved prefusion-closed trimers.bioRxiv [Preprint]. 2024 Mar 8:2024.03.07.583986. doi: 10.1101/2024.03.07.583986. bioRxiv. 2024. Update in: Nat Commun. 2024 Nov 16;15(1):9939. doi: 10.1038/s41467-024-54287-x PMID: 38496645 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
