

Molecular mechanism for activation of the 26S proteasome by ZFAND5
- PMID: 37595557
- PMCID: PMC10523585
- DOI: 10.1016/j.molcel.2023.07.023
Molecular mechanism for activation of the 26S proteasome by ZFAND5
Abstract
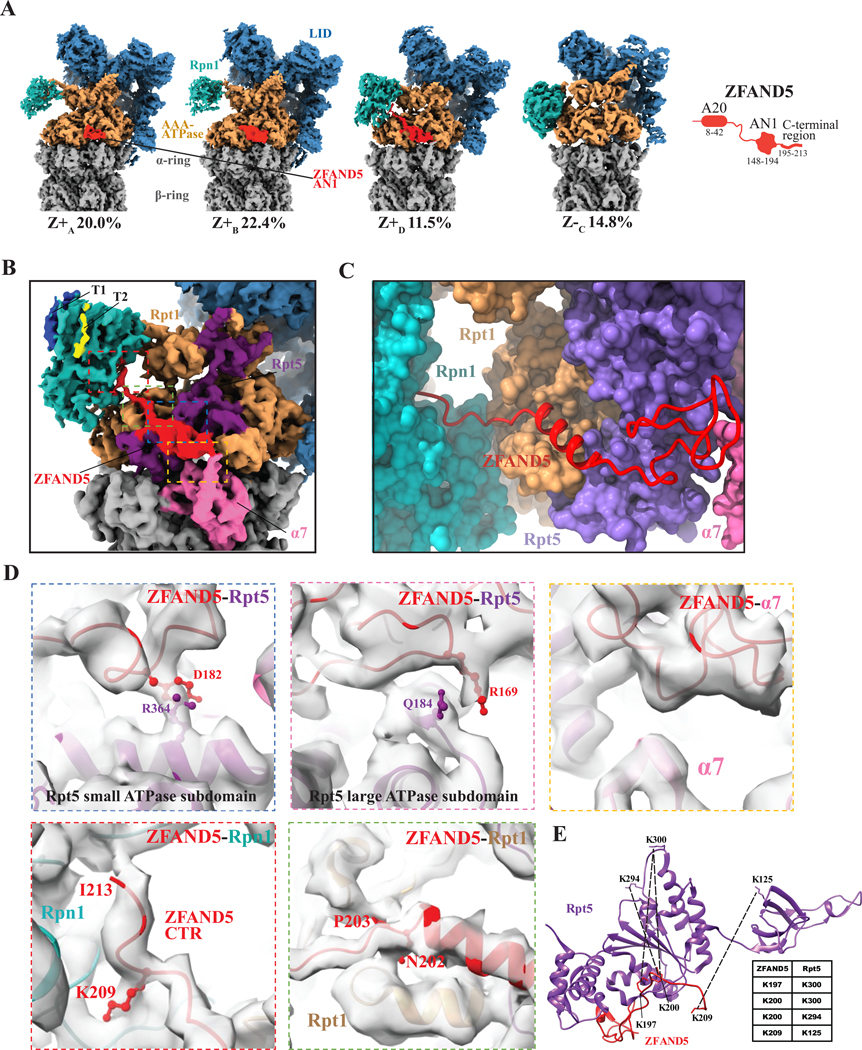
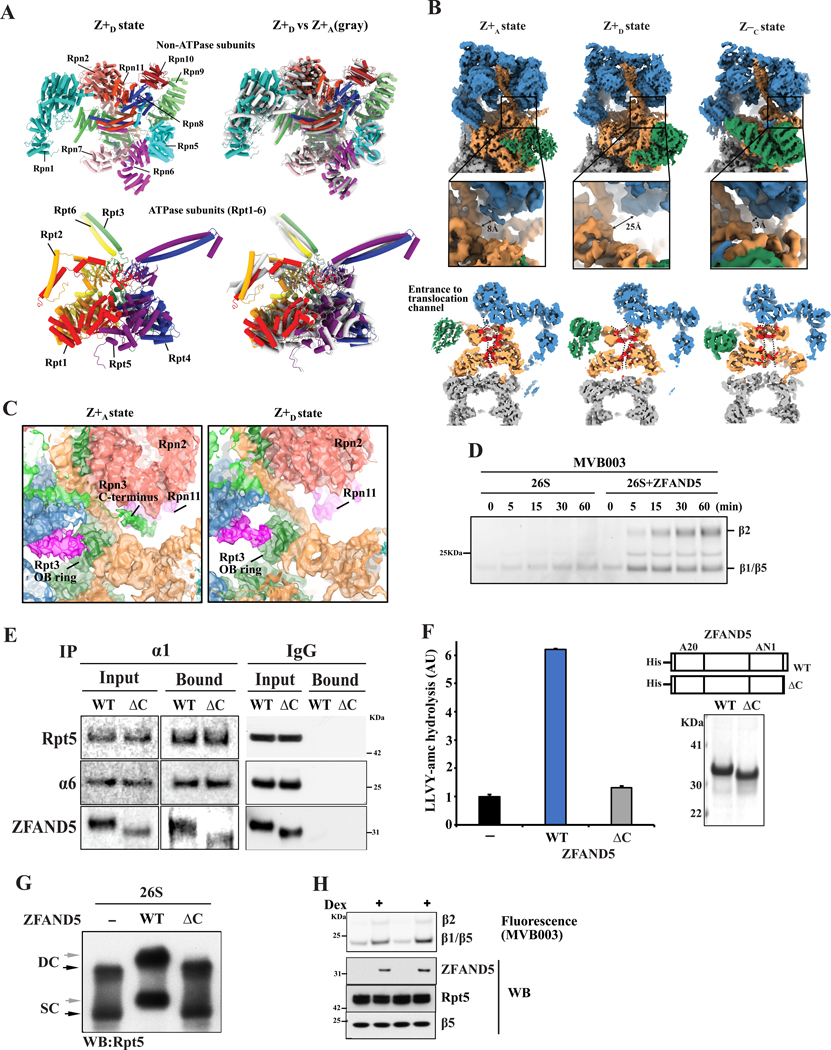
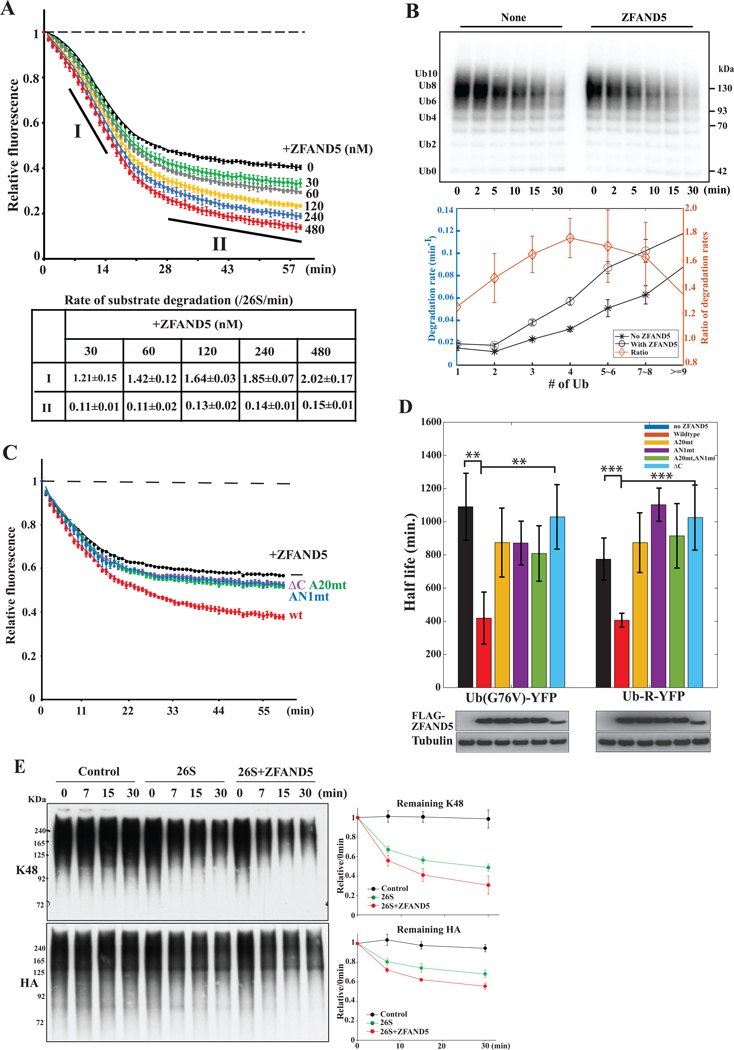
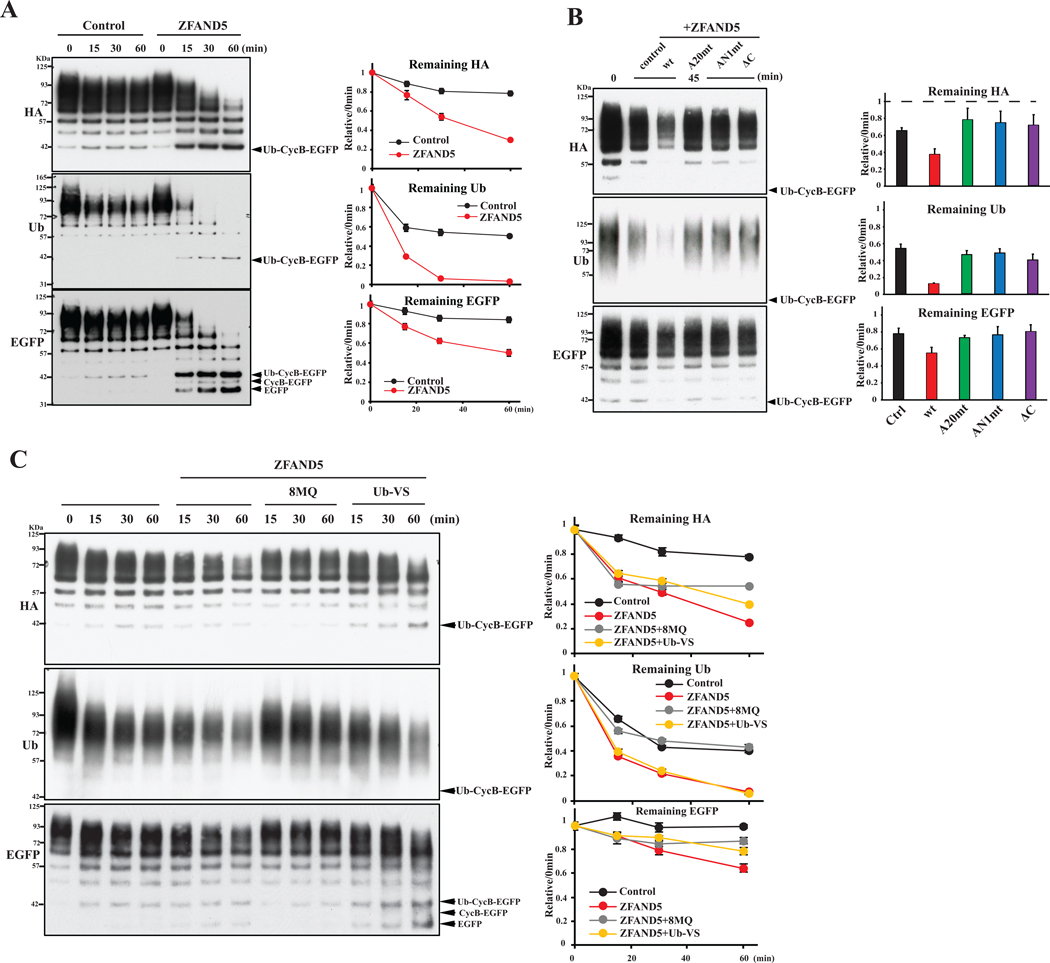
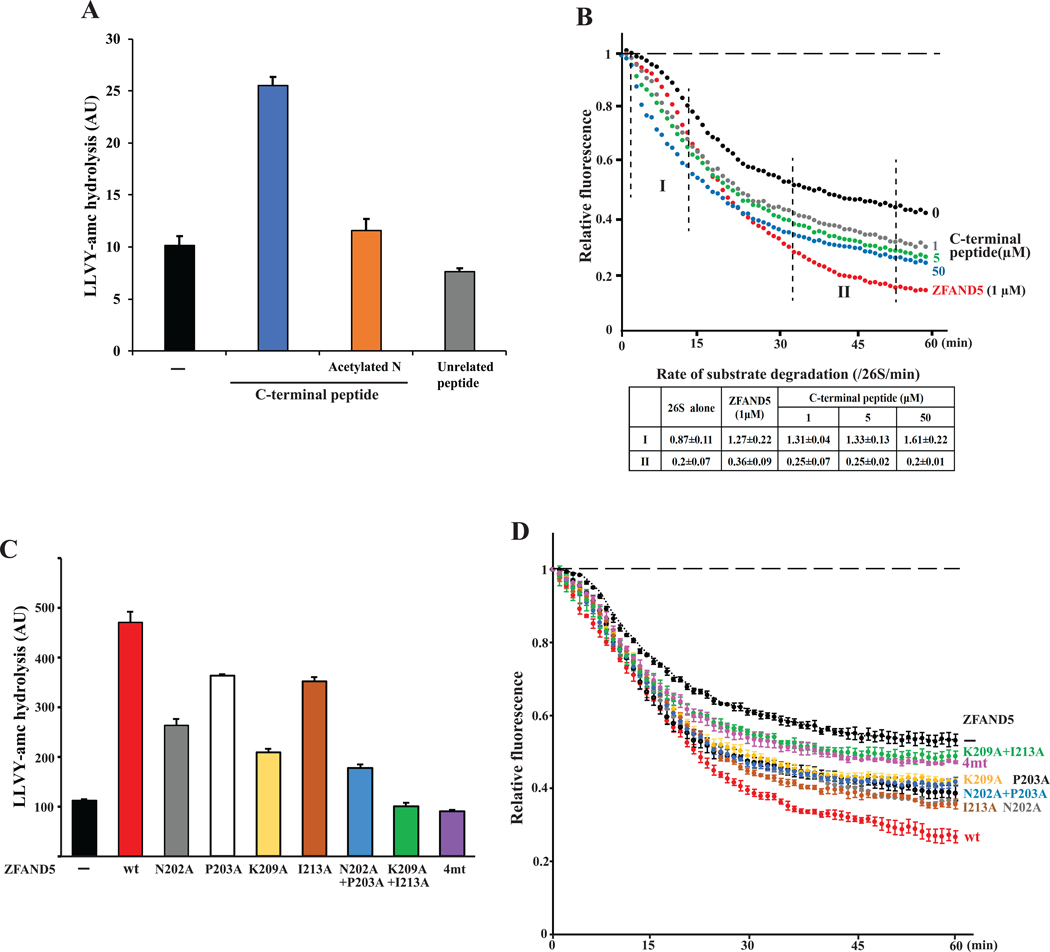
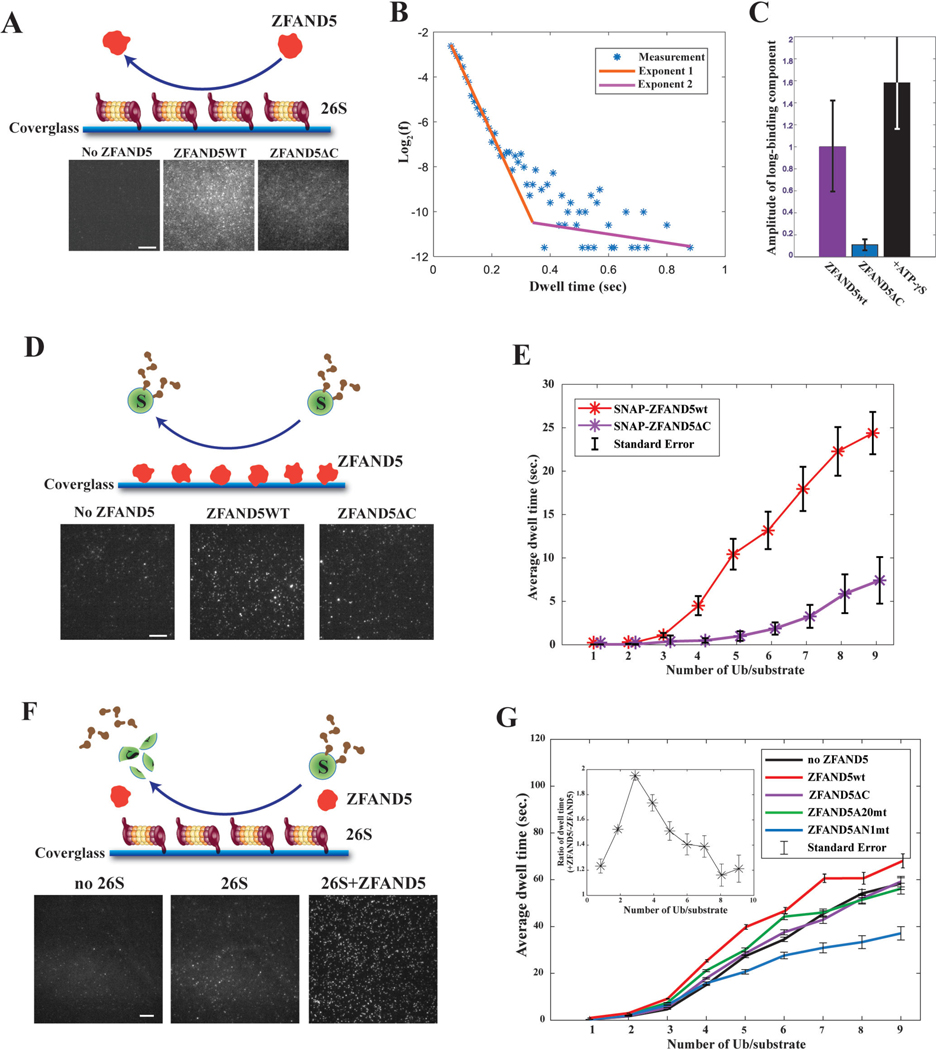
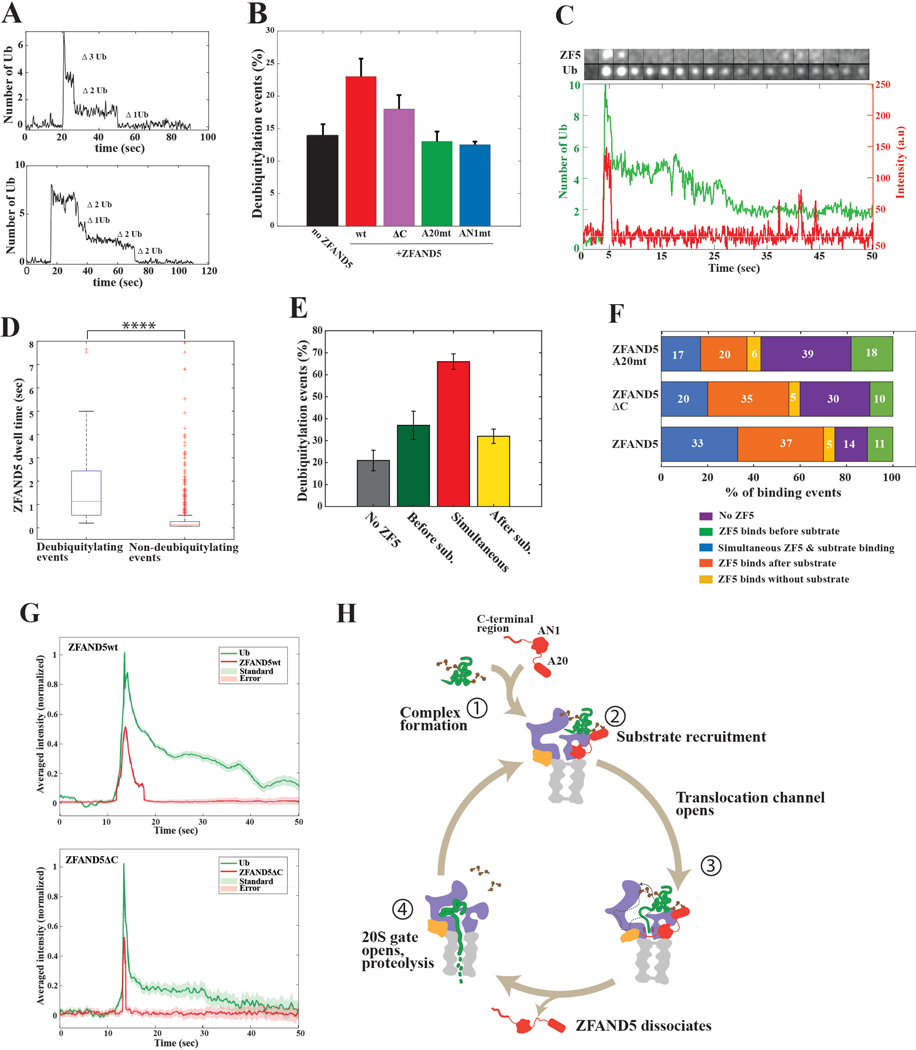
Various hormones, kinases, and stressors (fasting, heat shock) stimulate 26S proteasome activity. To understand how its capacity to degrade ubiquitylated proteins can increase, we studied mouse ZFAND5, which promotes protein degradation during muscle atrophy. Cryo-electron microscopy showed that ZFAND5 induces large conformational changes in the 19S regulatory particle. ZFAND5's AN1 Zn-finger domain interacts with the Rpt5 ATPase and its C terminus with Rpt1 ATPase and Rpn1, a ubiquitin-binding subunit. Upon proteasome binding, ZFAND5 widens the entrance of the substrate translocation channel, yet it associates only transiently with the proteasome. Dissociation of ZFAND5 then stimulates opening of the 20S proteasome gate. Using single-molecule microscopy, we showed that ZFAND5 binds ubiquitylated substrates, prolongs their association with proteasomes, and increases the likelihood that bound substrates undergo degradation, even though ZFAND5 dissociates before substrate deubiquitylation. These changes in proteasome conformation and reaction cycle can explain the accelerated degradation and suggest how other proteasome activators may stimulate proteolysis.
Keywords: ZFAND5; muscle atrophy; proteasome activation; proteasomes; protein degradation; ubiquitin.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors claim no competing interests.
Figures
Update of
-
Molecular mechanisms for activation of the 26S proteasome.bioRxiv [Preprint]. 2023 May 10:2023.05.09.540094. doi: 10.1101/2023.05.09.540094. bioRxiv. 2023. Update in: Mol Cell. 2023 Aug 17;83(16):2959-2975.e7. doi: 10.1016/j.molcel.2023.07.023. PMID: 37214989 Free PMC article. Updated. Preprint.
Similar articles
-
Molecular mechanisms for activation of the 26S proteasome.bioRxiv [Preprint]. 2023 May 10:2023.05.09.540094. doi: 10.1101/2023.05.09.540094. bioRxiv. 2023. Update in: Mol Cell. 2023 Aug 17;83(16):2959-2975.e7. doi: 10.1016/j.molcel.2023.07.023. PMID: 37214989 Free PMC article. Updated. Preprint.
-
ZFAND5/ZNF216 is an activator of the 26S proteasome that stimulates overall protein degradation.Proc Natl Acad Sci U S A. 2018 Oct 9;115(41):E9550-E9559. doi: 10.1073/pnas.1809934115. Epub 2018 Sep 25. Proc Natl Acad Sci U S A. 2018. PMID: 30254168 Free PMC article.
-
Mechanisms That Activate 26S Proteasomes and Enhance Protein Degradation.Biomolecules. 2021 May 22;11(6):779. doi: 10.3390/biom11060779. Biomolecules. 2021. PMID: 34067263 Free PMC article. Review.
-
USP14-regulated allostery of the human proteasome by time-resolved cryo-EM.Nature. 2022 May;605(7910):567-574. doi: 10.1038/s41586-022-04671-8. Epub 2022 Apr 27. Nature. 2022. PMID: 35477760 Free PMC article.
-
Structural Insights into Substrate Recognition and Processing by the 20S Proteasome.Biomolecules. 2021 Jan 24;11(2):148. doi: 10.3390/biom11020148. Biomolecules. 2021. PMID: 33498876 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
