

Identification of Regulatory Molecular "Hot Spots" for LH/PLOD Collagen Glycosyltransferase Activity
- PMID: 37446392
- PMCID: PMC10342707
- DOI: 10.3390/ijms241311213
Identification of Regulatory Molecular "Hot Spots" for LH/PLOD Collagen Glycosyltransferase Activity
Abstract
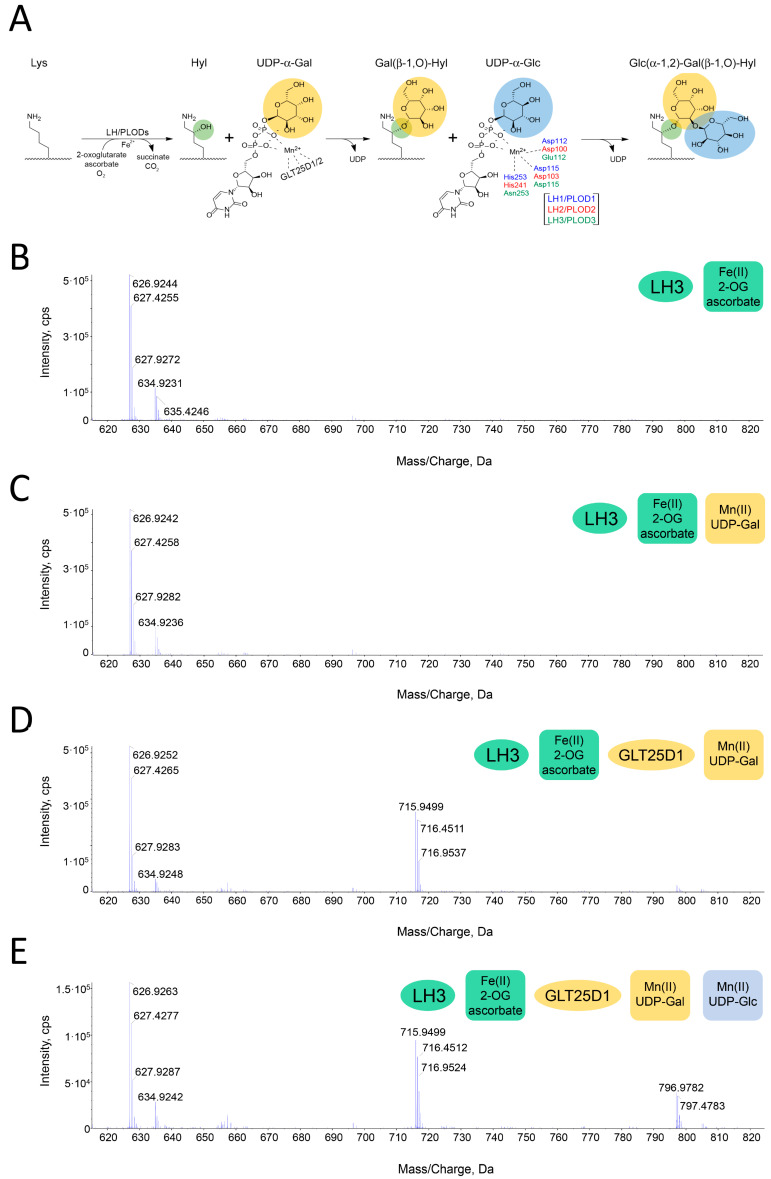
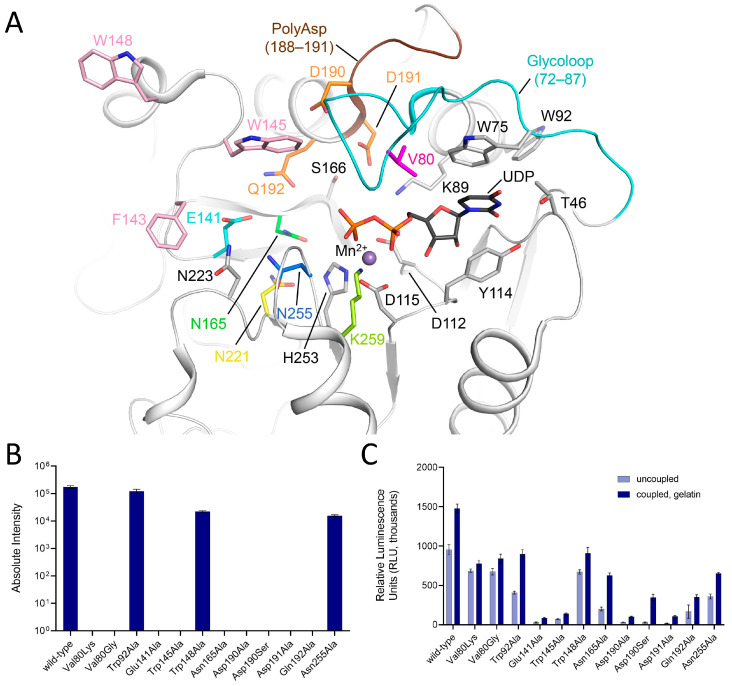
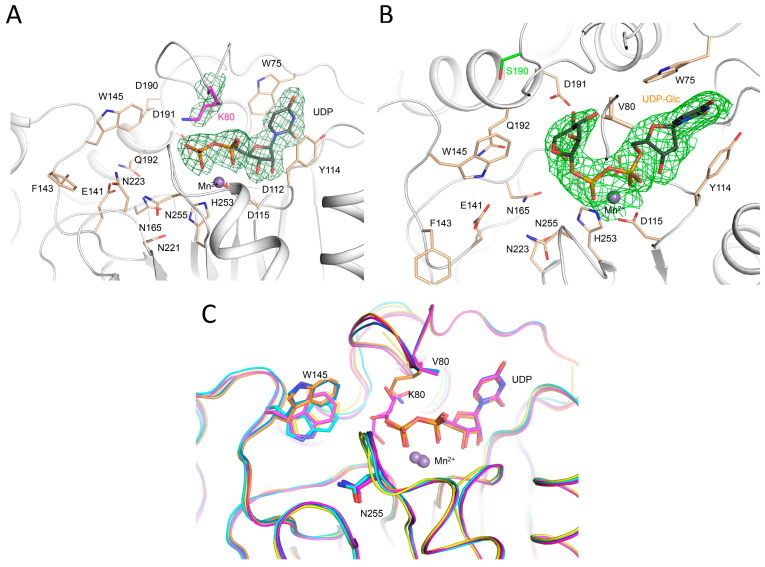
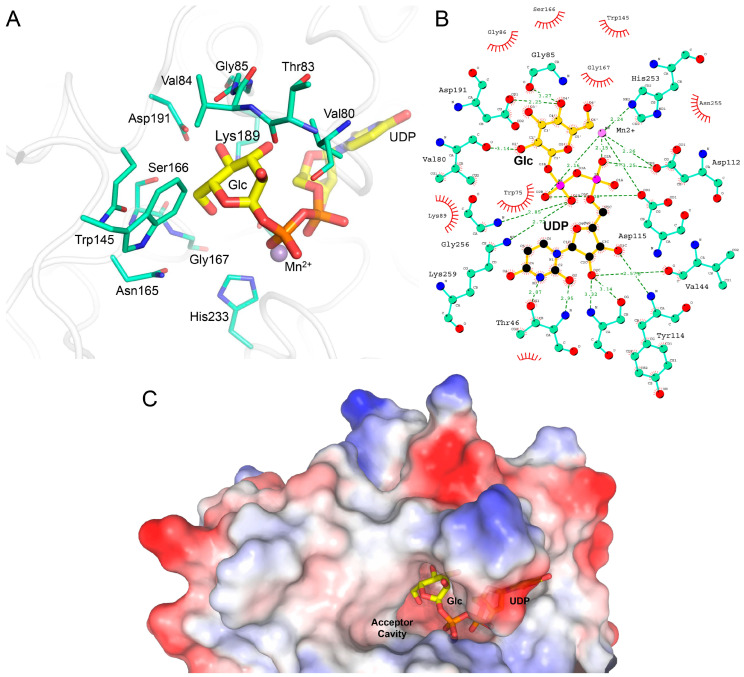
Hydroxylysine glycosylations are post-translational modifications (PTMs) essential for the maturation and homeostasis of fibrillar and non-fibrillar collagen molecules. The multifunctional collagen lysyl hydroxylase 3 (LH3/PLOD3) and the collagen galactosyltransferase GLT25D1 are the human enzymes that have been identified as being responsible for the glycosylation of collagen lysines, although a precise description of the contribution of each enzyme to these essential PTMs has not yet been provided in the literature. LH3/PLOD3 is thought to be capable of performing two chemically distinct collagen glycosyltransferase reactions using the same catalytic site: an inverting beta-1,O-galactosylation of hydroxylysines (Gal-T) and a retaining alpha-1,2-glucosylation of galactosyl hydroxylysines (Glc-T). In this work, we have combined indirect luminescence-based assays with direct mass spectrometry-based assays and molecular structure studies to demonstrate that LH3/PLOD3 only has Glc-T activity and that GLT25D1 only has Gal-T activity. Structure-guided mutagenesis confirmed that the Glc-T activity is defined by key residues in the first-shell environment of the glycosyltransferase catalytic site as well as by long-range contributions from residues within the same glycosyltransferase (GT) domain. By solving the molecular structures and characterizing the interactions and solving the molecular structures of human LH3/PLOD3 in complex with different UDP-sugar analogs, we show how these studies could provide insights for LH3/PLOD3 glycosyltransferase inhibitor development. Collectively, our data provide new tools for the direct investigation of collagen hydroxylysine PTMs and a comprehensive overview of the complex network of shapes, charges, and interactions that enable LH3/PLOD3 glycosyltransferase activities, expanding the molecular framework and facilitating an improved understanding and manipulation of glycosyltransferase functions in biomedical applications.
Keywords: collagen biosynthesis; collagen glycosylations; extracellular matrix; glycosyltransferase; lysyl hydroxylase; post-translational modifications.
Conflict of interest statement
Authors declare no conflict of interest.
Figures
Similar articles
-
Expanding the lysyl hydroxylase toolbox: new insights into the localization and activities of lysyl hydroxylase 3 (LH3).J Cell Physiol. 2007 Aug;212(2):323-9. doi: 10.1002/jcp.21036. J Cell Physiol. 2007. PMID: 17516569 Review.
-
Molecular architecture of the multifunctional collagen lysyl hydroxylase and glycosyltransferase LH3.Nat Commun. 2018 Aug 8;9(1):3163. doi: 10.1038/s41467-018-05631-5. Nat Commun. 2018. PMID: 30089812 Free PMC article.
-
Collagen hydroxylysine glycosylation: non-conventional substrates for atypical glycosyltransferase enzymes.Biochem Soc Trans. 2021 Apr 30;49(2):855-866. doi: 10.1042/BST20200767. Biochem Soc Trans. 2021. PMID: 33704379 Review.
-
The glycosyltransferase activities of lysyl hydroxylase 3 (LH3) in the extracellular space are important for cell growth and viability.J Cell Mol Med. 2009 Mar;13(3):508-21. doi: 10.1111/j.1582-4934.2008.00286.x. J Cell Mol Med. 2009. PMID: 18298658 Free PMC article.
-
Lysyl hydroxylase 3 glucosylates galactosylhydroxylysine residues in type I collagen in osteoblast culture.J Biol Chem. 2011 Mar 18;286(11):8846-56. doi: 10.1074/jbc.M110.178509. Epub 2011 Jan 10. J Biol Chem. 2011. PMID: 21220425 Free PMC article.
Cited by
-
Glycosylation Modulates the Structure and Functions of Collagen: A Review.Molecules. 2024 Mar 22;29(7):1417. doi: 10.3390/molecules29071417. Molecules. 2024. PMID: 38611696 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
