

Retrotransposons hijack alt-EJ for DNA replication and eccDNA biogenesis
- PMID: 37438532
- PMCID: PMC10691919
- DOI: 10.1038/s41586-023-06327-7
Retrotransposons hijack alt-EJ for DNA replication and eccDNA biogenesis
Abstract
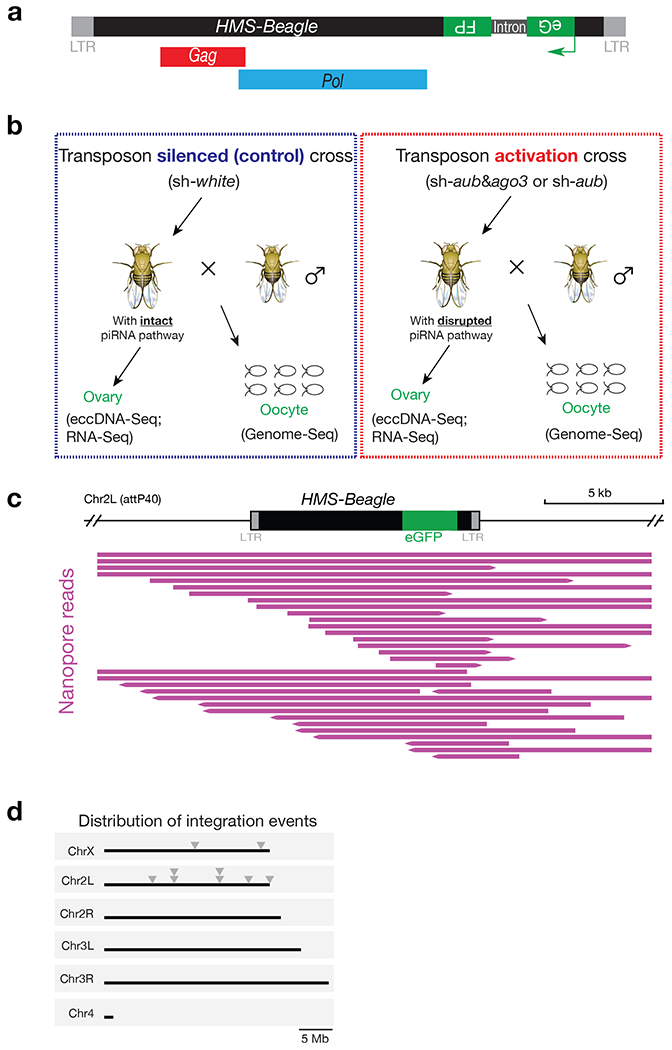
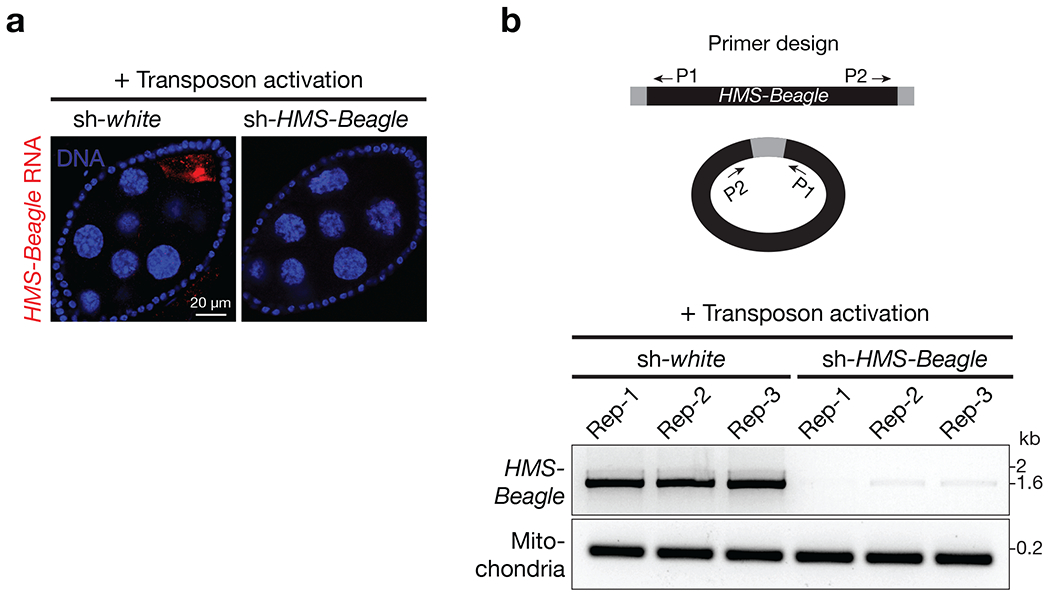
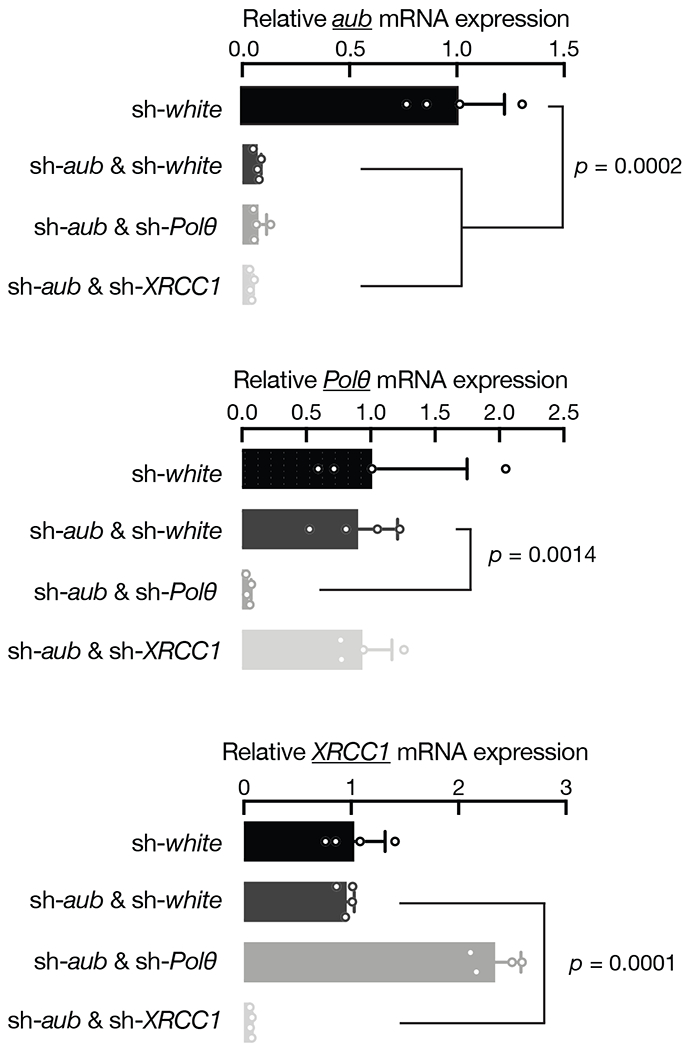
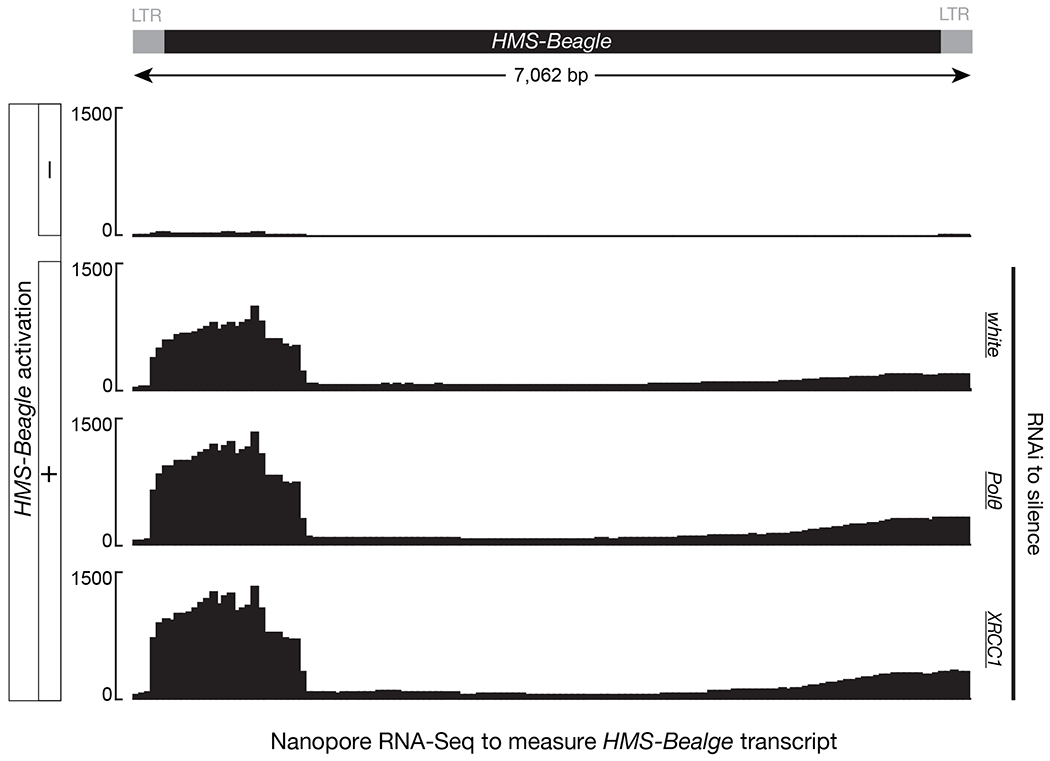
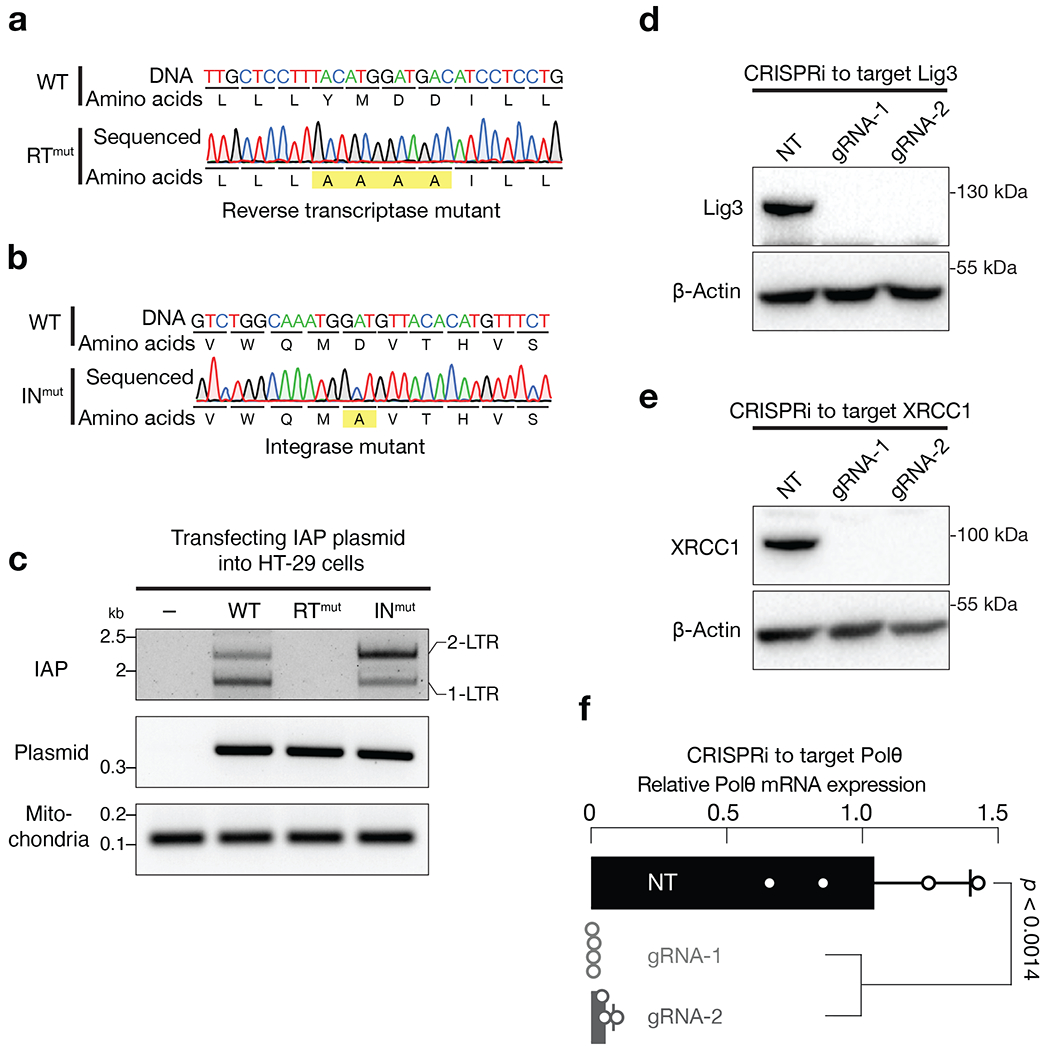
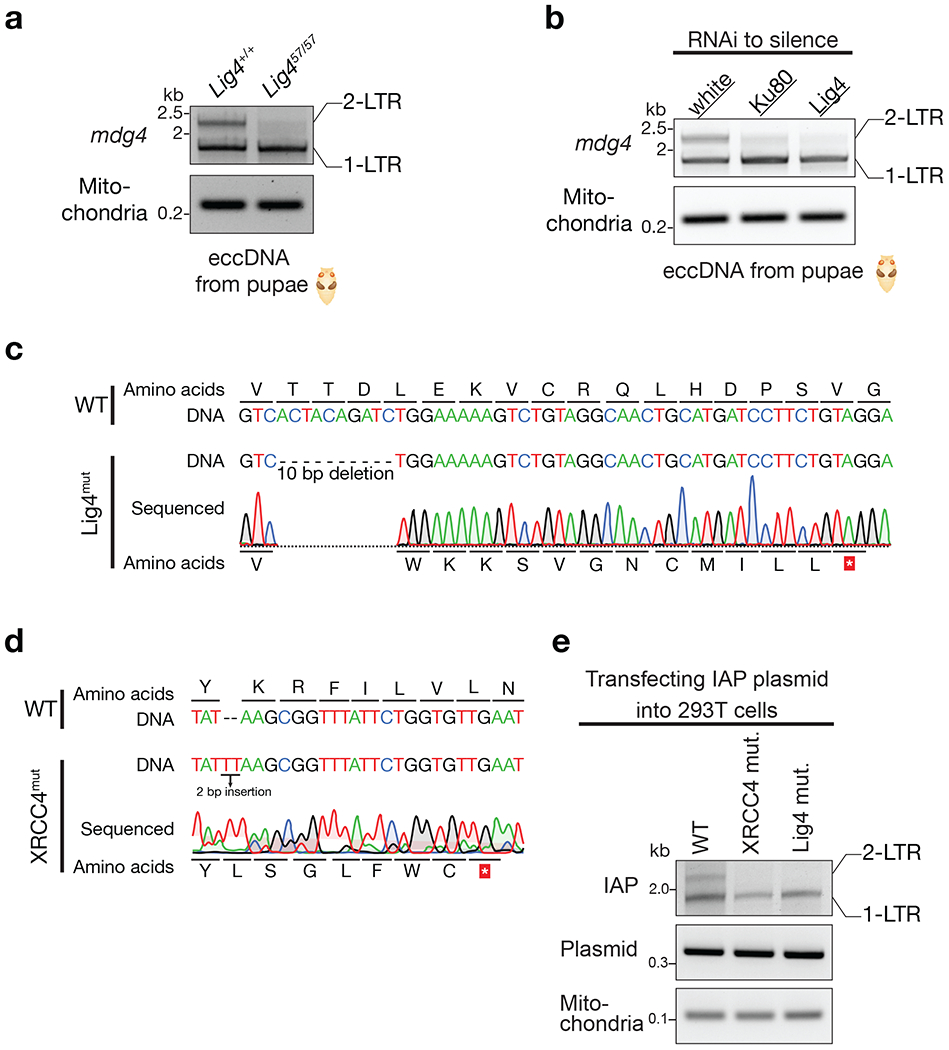
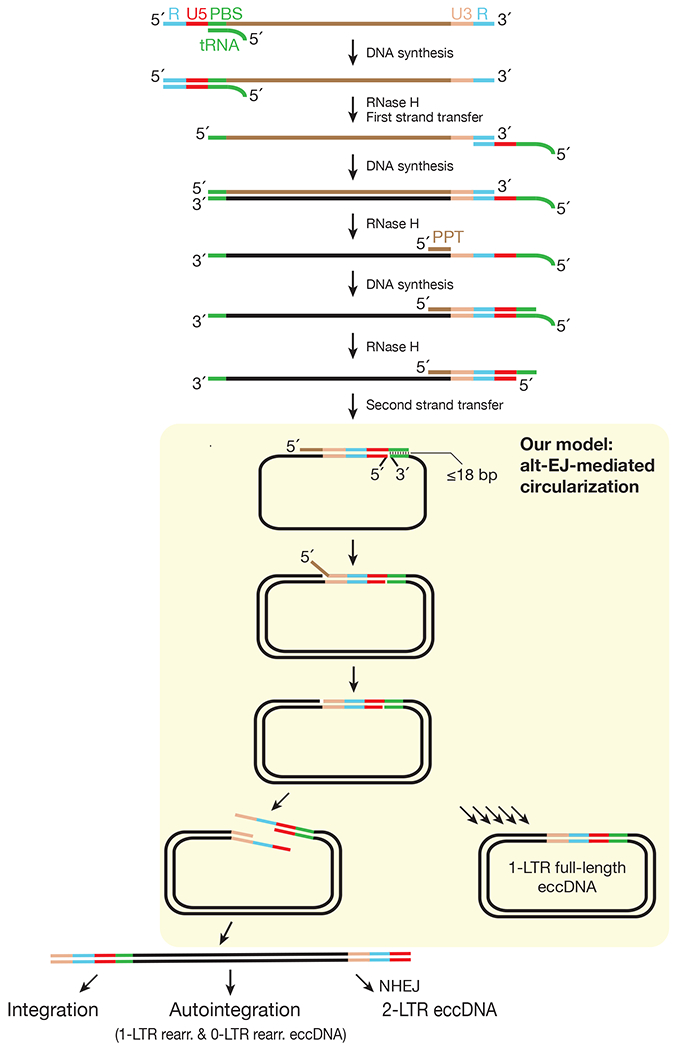
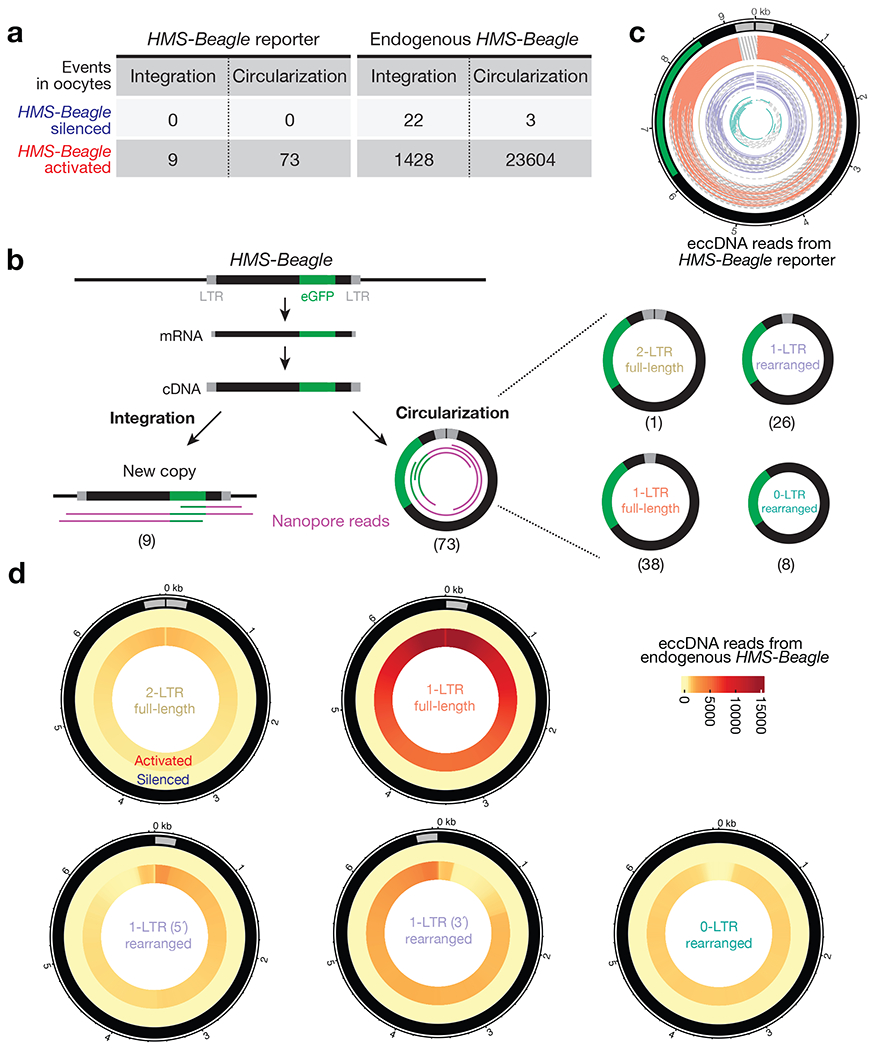
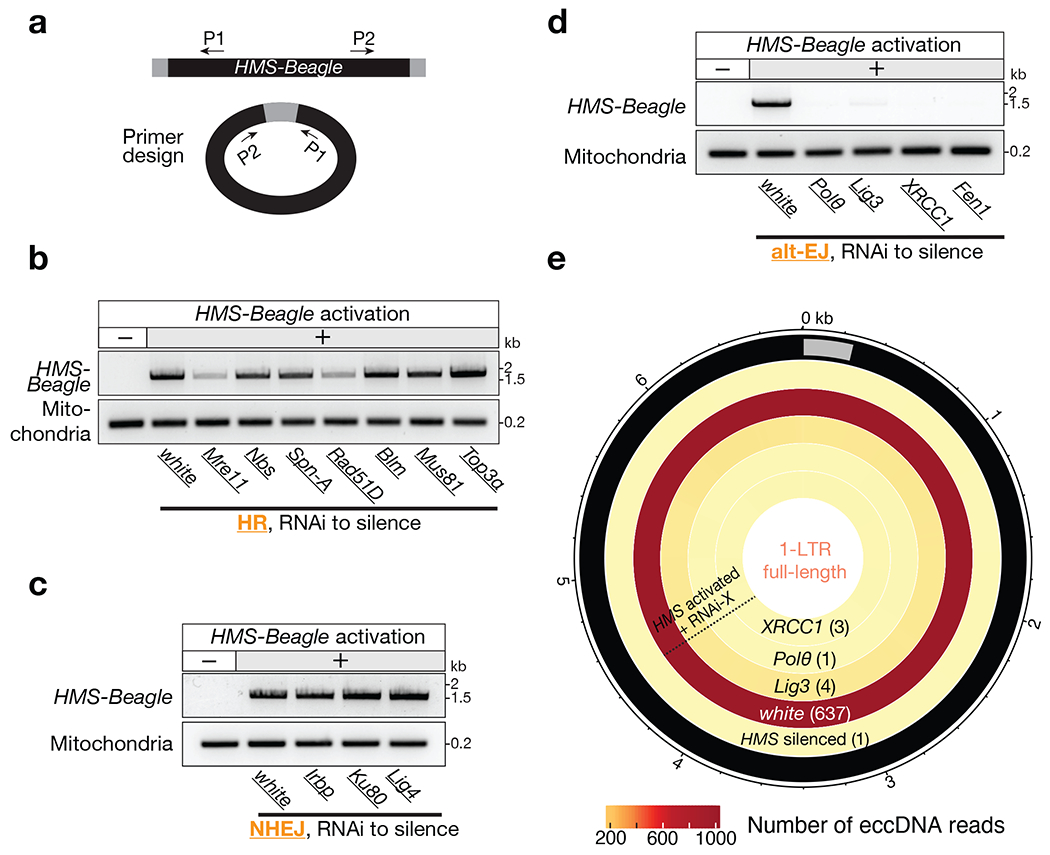
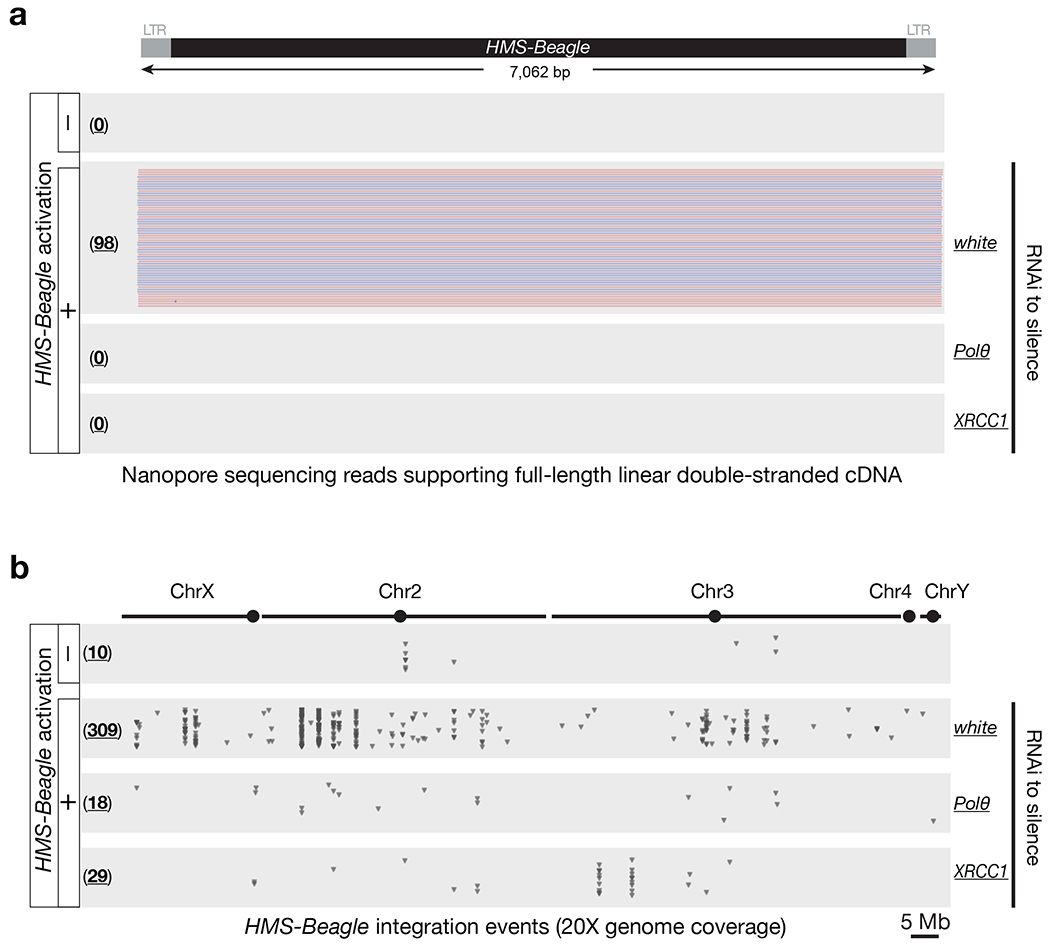
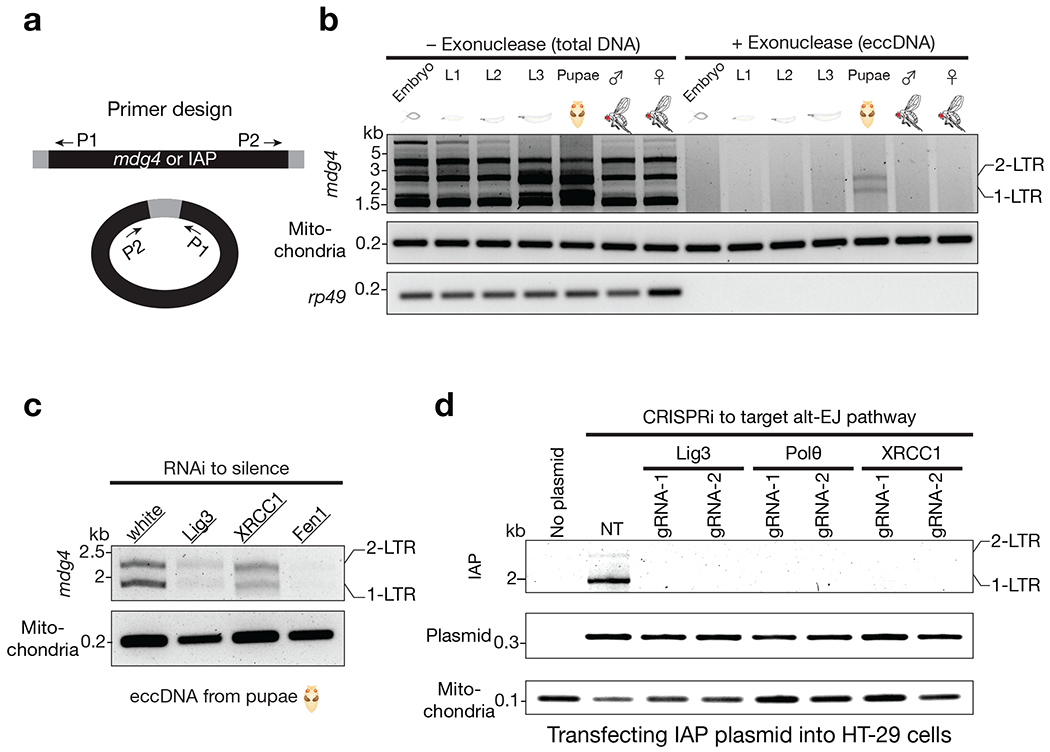
Retrotransposons are highly enriched in the animal genome1-3. The activation of retrotransposons can rewrite host DNA information and fundamentally impact host biology1-3. Although developmental activation of retrotransposons can offer benefits for the host, such as against virus infection, uncontrolled activation promotes disease or potentially drives ageing1-5. After activation, retrotransposons use their mRNA as templates to synthesize double-stranded DNA for making new insertions in the host genome1-3,6. Although the reverse transcriptase that they encode can synthesize the first-strand DNA1-3,6, how the second-strand DNA is generated remains largely unclear. Here we report that retrotransposons hijack the alternative end-joining (alt-EJ) DNA repair process of the host for a circularization step to synthesize their second-strand DNA. We used Nanopore sequencing to examine the fates of replicated retrotransposon DNA, and found that 10% of them achieve new insertions, whereas 90% exist as extrachromosomal circular DNA (eccDNA). Using eccDNA production as a readout, further genetic screens identified factors from alt-EJ as essential for retrotransposon replication. alt-EJ drives the second-strand synthesis of the long terminal repeat retrotransposon DNA through a circularization process and is therefore necessary for eccDNA production and new insertions. Together, our study reveals that alt-EJ is essential in driving the propagation of parasitic genomic retroelements. Our study uncovers a conserved function of this understudied DNA repair process, and provides a new perspective to understand-and potentially control-the retrotransposon life cycle.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
COMPETING INTERESTS
Z.Z, F.Y., and W.S. are co-inventors on a US provisional patent application filed by Duke University related to this work.
Figures




Similar articles
-
DNA-directed DNA polymerase and strand displacement activity of the reverse transcriptase encoded by the R2 retrotransposon.J Mol Biol. 2007 Nov 23;374(2):322-33. doi: 10.1016/j.jmb.2007.09.047. Epub 2007 Sep 20. J Mol Biol. 2007. PMID: 17936300 Free PMC article.
-
The reverse transcriptase of the R2 non-LTR retrotransposon: continuous synthesis of cDNA on non-continuous RNA templates.J Mol Biol. 2002 Feb 22;316(3):459-73. doi: 10.1006/jmbi.2001.5369. J Mol Biol. 2002. PMID: 11866511
-
Sequencing the extrachromosomal circular mobilome reveals retrotransposon activity in plants.PLoS Genet. 2017 Feb 17;13(2):e1006630. doi: 10.1371/journal.pgen.1006630. eCollection 2017 Feb. PLoS Genet. 2017. PMID: 28212378 Free PMC article.
-
Alternative end-joining repair pathways are the ultimate backup for abrogated classical non-homologous end-joining and homologous recombination repair: Implications for the formation of chromosome translocations.Mutat Res Genet Toxicol Environ Mutagen. 2015 Nov;793:166-75. doi: 10.1016/j.mrgentox.2015.07.001. Epub 2015 Jul 4. Mutat Res Genet Toxicol Environ Mutagen. 2015. PMID: 26520387 Review.
-
"In the beginning": initiation of minus strand DNA synthesis in retroviruses and LTR-containing retrotransposons.Biochemistry. 2003 Dec 16;42(49):14349-55. doi: 10.1021/bi030201q. Biochemistry. 2003. PMID: 14661945 Review.
Cited by
-
The Molecular Impacts of Retrotransposons in Development and Diseases.Int J Mol Sci. 2023 Nov 16;24(22):16418. doi: 10.3390/ijms242216418. Int J Mol Sci. 2023. PMID: 38003607 Free PMC article. Review.
-
Retrotransposons and Diabetes Mellitus.Epigenomes. 2024 Sep 6;8(3):35. doi: 10.3390/epigenomes8030035. Epigenomes. 2024. PMID: 39311137 Free PMC article. Review.
-
Meeting report: transposable elements at the crossroads of evolution, health and disease 2023.Mob DNA. 2023 Nov 27;14(1):19. doi: 10.1186/s13100-023-00307-4. Mob DNA. 2023. PMID: 38012685 Free PMC article.
-
Comparative analysis of methodologies for detecting extrachromosomal circular DNA.Nat Commun. 2024 Oct 25;15(1):9208. doi: 10.1038/s41467-024-53496-8. Nat Commun. 2024. PMID: 39448595 Free PMC article.
-
Circle-seq based method for eccDNA synthesis and its application as a canonical promoter independent vector for robust microRNA overexpression.Comput Struct Biotechnol J. 2023 Dec 20;23:358-368. doi: 10.1016/j.csbj.2023.12.019. eCollection 2024 Dec. Comput Struct Biotechnol J. 2023. PMID: 38223344 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
