

A gain-of-function variation in PLCG1 causes a new immune dysregulation disease
- PMID: 37422272
- PMCID: PMC10770301
- DOI: 10.1016/j.jaci.2023.06.020
A gain-of-function variation in PLCG1 causes a new immune dysregulation disease
Abstract
Background: Phospholipase C (PLC) γ1 is a critical enzyme regulating nuclear factor-κB (NF-κB), extracellular signal-related kinase, mitogen-activated protein kinase, and nuclear factor of activated T cells signaling pathways, yet germline PLCG1 mutation in human disease has not been reported.
Objective: We aimed to investigate the molecular pathogenesis of a PLCG1 activating variant in a patient with immune dysregulation.
Methods: Whole exome sequencing was used to identify the patient's pathogenic variants. Bulk RNA sequencing, single-cell RNA sequencing, quantitative PCR, cytometry by time of flight, immunoblotting, flow cytometry, luciferase assay, IP-One ELISA, calcium flux assay, and cytokine measurements in patient PBMCs and T cells and COS-7 and Jurkat cell lines were used to define inflammatory signatures and assess the impact of the PLCG1 variant on protein function and immune signaling.
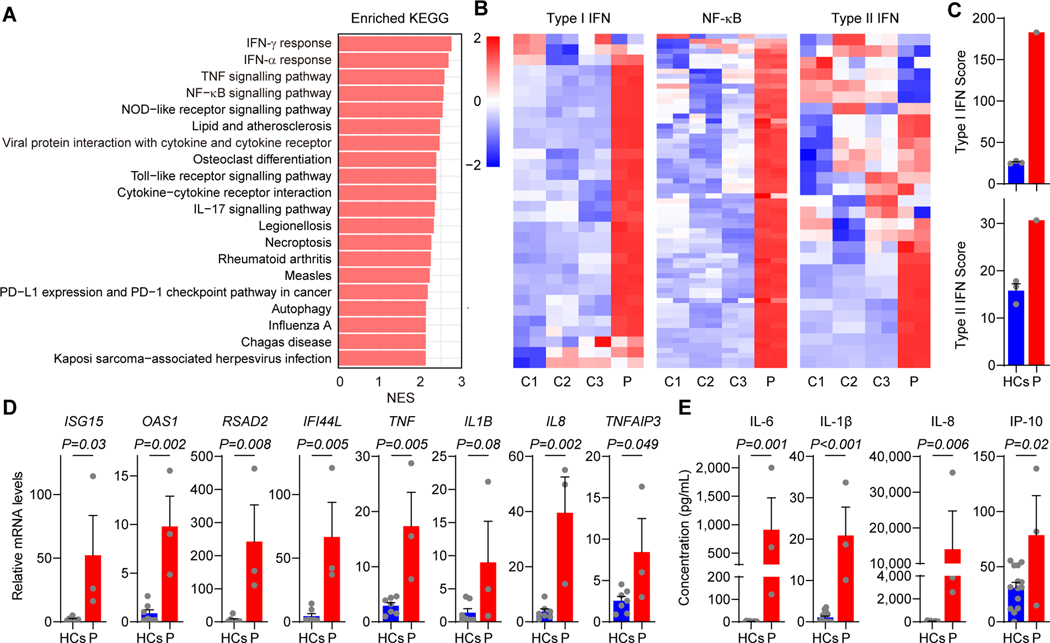
Results: We identified a novel and de novo heterozygous PLCG1 variant, p.S1021F, in a patient presenting with early-onset immune dysregulation disease. We demonstrated that the S1021F variant is a gain-of-function variant, leading to increased inositol-1,4,5-trisphosphate production, intracellular Ca2+ release, and increased phosphorylation of extracellular signal-related kinase, p65, and p38. The transcriptome and protein expression at the single-cell level revealed exacerbated inflammatory responses in the patient's T cells and monocytes. The PLCG1 activating variant resulted in enhanced NF-κB and type II interferon pathways in T cells, and hyperactivated NF-κB and type I interferon pathways in monocytes. Treatment with either PLCγ1 inhibitor or Janus kinase inhibitor reversed the upregulated gene expression profile in vitro.
Conclusions: Our study highlights the critical role of PLCγ1 in maintaining immune homeostasis. We illustrate immune dysregulation as a consequence of PLCγ1 activation and provide insight into therapeutic targeting of PLCγ1.
Keywords: PLCG1; autoinflammation; gain-of-function variant; immune dysregulation.
Copyright © 2023 American Academy of Allergy, Asthma & Immunology. All rights reserved.
Conflict of interest statement
Figures



Similar articles
-
Characterization of a Novel Pathogenic PLCG2 Variant Leading to APLAID Syndrome Responsive to a TNF Inhibitor.Arthritis Rheumatol. 2024 Nov;76(11):1670-1678. doi: 10.1002/art.42948. Epub 2024 Aug 6. Arthritis Rheumatol. 2024. PMID: 38965708
-
Frequent and Persistent PLCG1 Mutations in Sézary Cells Directly Enhance PLCγ1 Activity and Stimulate NFκB, AP-1, and NFAT Signaling.J Invest Dermatol. 2020 Feb;140(2):380-389.e4. doi: 10.1016/j.jid.2019.07.693. Epub 2019 Jul 31. J Invest Dermatol. 2020. PMID: 31376383
-
Analysis of platelet-derived growth factor-induced phospholipase D activation in mouse embryo fibroblasts lacking phospholipase C-gamma1.J Biol Chem. 1998 Aug 7;273(32):20517-24. doi: 10.1074/jbc.273.32.20517. J Biol Chem. 1998. PMID: 9685408
-
Reprint of Neutrophil cell surface receptors and their intracellular signal transduction pathways.Int Immunopharmacol. 2013 Dec;17(4):1185-97. doi: 10.1016/j.intimp.2013.11.010. Epub 2013 Nov 18. Int Immunopharmacol. 2013. PMID: 24263067 Review.
-
Neutrophil cell surface receptors and their intracellular signal transduction pathways.Int Immunopharmacol. 2013 Nov;17(3):638-50. doi: 10.1016/j.intimp.2013.06.034. Epub 2013 Aug 30. Int Immunopharmacol. 2013. PMID: 23994464 Free PMC article. Review.
Cited by
-
A novel micellular fluorogenic substrate for quantitating the activity of 1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase gamma (PLCγ) enzymes.PLoS One. 2024 Mar 29;19(3):e0299541. doi: 10.1371/journal.pone.0299541. eCollection 2024. PLoS One. 2024. PMID: 38551930 Free PMC article.
-
Nucleotide metabolism, leukodystrophies, and CNS pathology.J Inherit Metab Dis. 2024 Feb 29:10.1002/jimd.12721. doi: 10.1002/jimd.12721. Online ahead of print. J Inherit Metab Dis. 2024. PMID: 38421058
-
De novo variants in PLCG1 are associated with hearing impairment, ocular pathology, and cardiac defects.medRxiv [Preprint]. 2024 Jan 9:2024.01.08.23300523. doi: 10.1101/2024.01.08.23300523. medRxiv. 2024. PMID: 38260438 Free PMC article. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
