

Lateral membrane organization as target of an antimicrobial peptidomimetic compound
- PMID: 37419980
- PMCID: PMC10328936
- DOI: 10.1038/s41467-023-39726-5
Lateral membrane organization as target of an antimicrobial peptidomimetic compound
Abstract
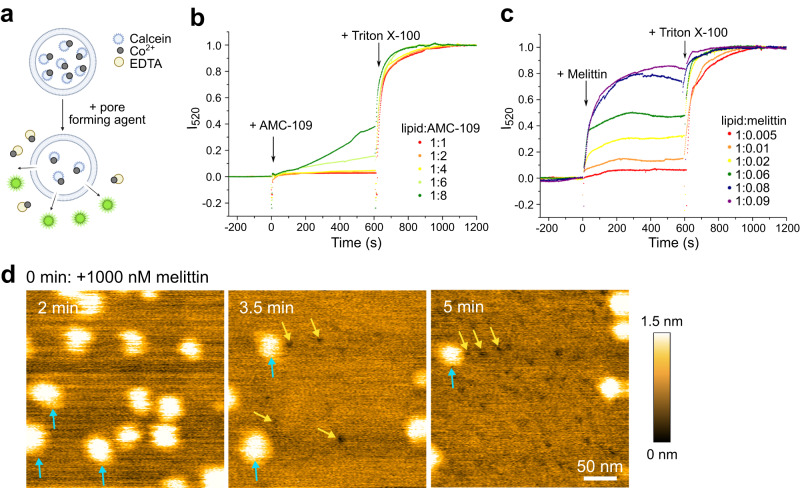
Antimicrobial resistance is one of the leading concerns in medical care. Here we study the mechanism of action of an antimicrobial cationic tripeptide, AMC-109, by combining high speed-atomic force microscopy, molecular dynamics, fluorescence assays, and lipidomic analysis. We show that AMC-109 activity on negatively charged membranes derived from Staphylococcus aureus consists of two crucial steps. First, AMC-109 self-assembles into stable aggregates consisting of a hydrophobic core and a cationic surface, with specificity for negatively charged membranes. Second, upon incorporation into the membrane, individual peptides insert into the outer monolayer, affecting lateral membrane organization and dissolving membrane nanodomains, without forming pores. We propose that membrane domain dissolution triggered by AMC-109 may affect crucial functions such as protein sorting and cell wall synthesis. Our results indicate that the AMC-109 mode of action resembles that of the disinfectant benzalkonium chloride (BAK), but with enhanced selectivity for bacterial membranes.
© 2023. The Author(s).
Conflict of interest statement
JSMS and WS are employed by Amicoat AS, the producer of AMC-109. The remaining authors declare no competing interests.
Figures
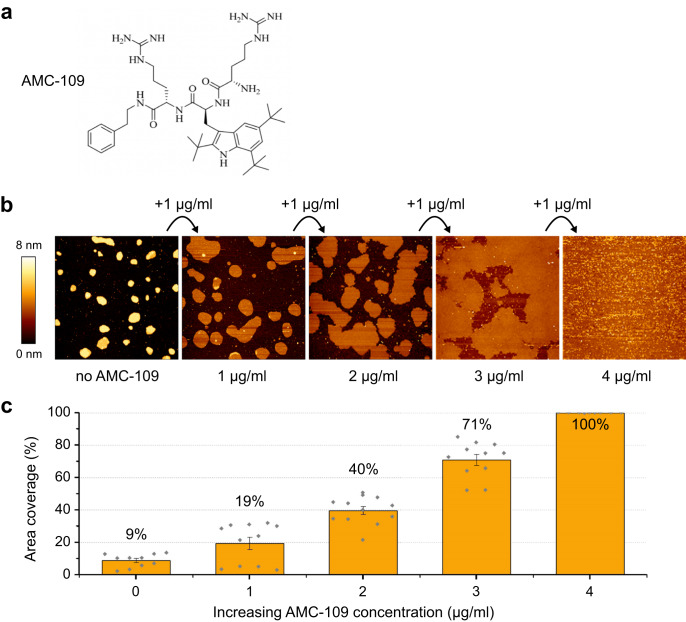
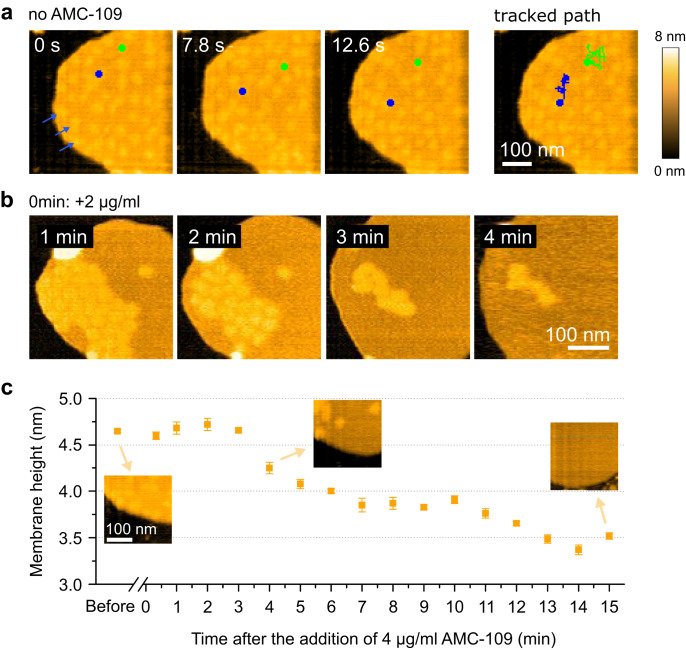
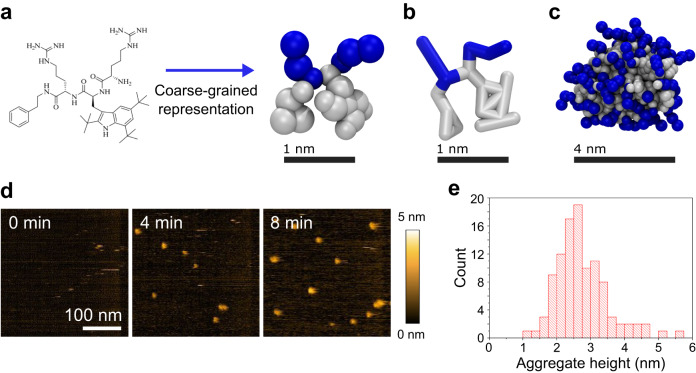
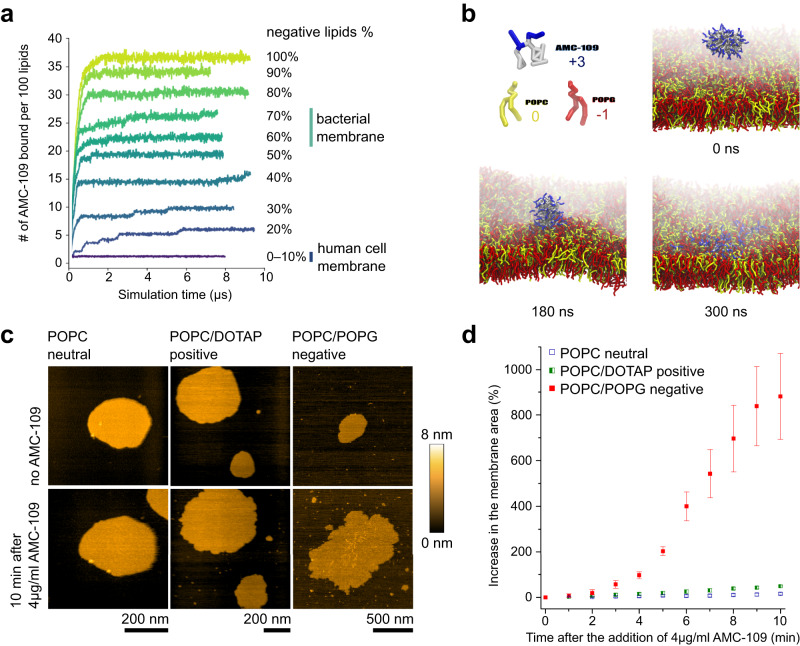

Similar articles
-
De novo design and synthesis of ultra-short peptidomimetic antibiotics having dual antimicrobial and anti-inflammatory activities.PLoS One. 2013 Nov 26;8(11):e80025. doi: 10.1371/journal.pone.0080025. eCollection 2013. PLoS One. 2013. PMID: 24302996 Free PMC article.
-
Norbornane-based cationic antimicrobial peptidomimetics targeting the bacterial membrane.Eur J Med Chem. 2018 Dec 5;160:9-22. doi: 10.1016/j.ejmech.2018.09.072. Epub 2018 Oct 3. Eur J Med Chem. 2018. PMID: 30316060 Free PMC article.
-
Amphiphilic cationic β(3R3)-peptides: membrane active peptidomimetics and their potential as antimicrobial agents.Biomacromolecules. 2014 May 12;15(5):1687-95. doi: 10.1021/bm500101w. Epub 2014 Apr 15. Biomacromolecules. 2014. PMID: 24694059
-
Use of antimicrobial peptides against microbial biofilms: advantages and limits.Curr Med Chem. 2011;18(2):256-79. doi: 10.2174/092986711794088399. Curr Med Chem. 2011. PMID: 21110801 Review.
-
Antimicrobial AApeptides.Curr Top Med Chem. 2017;17(11):1266-1279. doi: 10.2174/1568026616666161018145945. Curr Top Med Chem. 2017. PMID: 27758686 Free PMC article. Review.
Cited by
-
Cell-Free Systems: Ideal Platforms for Accelerating the Discovery and Production of Peptide-Based Antibiotics.Int J Mol Sci. 2024 Aug 22;25(16):9109. doi: 10.3390/ijms25169109. Int J Mol Sci. 2024. PMID: 39201795 Free PMC article. Review.
-
Recent Advances in Amphipathic Peptidomimetics as Antimicrobial Agents to Combat Drug Resistance.Molecules. 2024 May 24;29(11):2492. doi: 10.3390/molecules29112492. Molecules. 2024. PMID: 38893366 Free PMC article. Review.
-
Multifaceted Activity of Fabimycin: Insights from Molecular Dynamics Studies on Bacterial Membrane Models.J Chem Inf Model. 2024 May 27;64(10):4204-4217. doi: 10.1021/acs.jcim.4c00228. Epub 2024 May 11. J Chem Inf Model. 2024. PMID: 38733348 Free PMC article.
-
Membrane-Active Antibiotics Affect Domains in Bacterial Membranes as the First Step of Their Activity.Nano Lett. 2024 Sep 25;24(38):11800-11807. doi: 10.1021/acs.nanolett.4c01873. Epub 2024 Aug 15. Nano Lett. 2024. PMID: 39145544 Free PMC article.
-
Preventing E. coli Biofilm Formation with Antimicrobial Peptide-Functionalized Surface Coatings: Recognizing the Dependence on the Bacterial Binding Mode Using Live-Cell Microscopy.ACS Appl Mater Interfaces. 2024 Feb 14;16(6):6799-6812. doi: 10.1021/acsami.3c16004. Epub 2024 Jan 31. ACS Appl Mater Interfaces. 2024. PMID: 38294883 Free PMC article.
References
-
- Dosler S. Antimicrobial peptides: Coming to the end of antibiotic era, the most promising agents. Istanbul J. Pharm. 2017;47:72–76. doi: 10.5152/IstanbulJPharm.2017.0012. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
