

This is a preprint.
Structural Organization of the Retriever-CCC Endosomal Recycling Complex
- PMID: 37333304
- PMCID: PMC10274727
- DOI: 10.1101/2023.06.06.543888
Structural Organization of the Retriever-CCC Endosomal Recycling Complex
Update in
-
Structural organization of the retriever-CCC endosomal recycling complex.Nat Struct Mol Biol. 2024 Jun;31(6):910-924. doi: 10.1038/s41594-023-01184-4. Epub 2023 Dec 7. Nat Struct Mol Biol. 2024. PMID: 38062209 Free PMC article.
Abstract
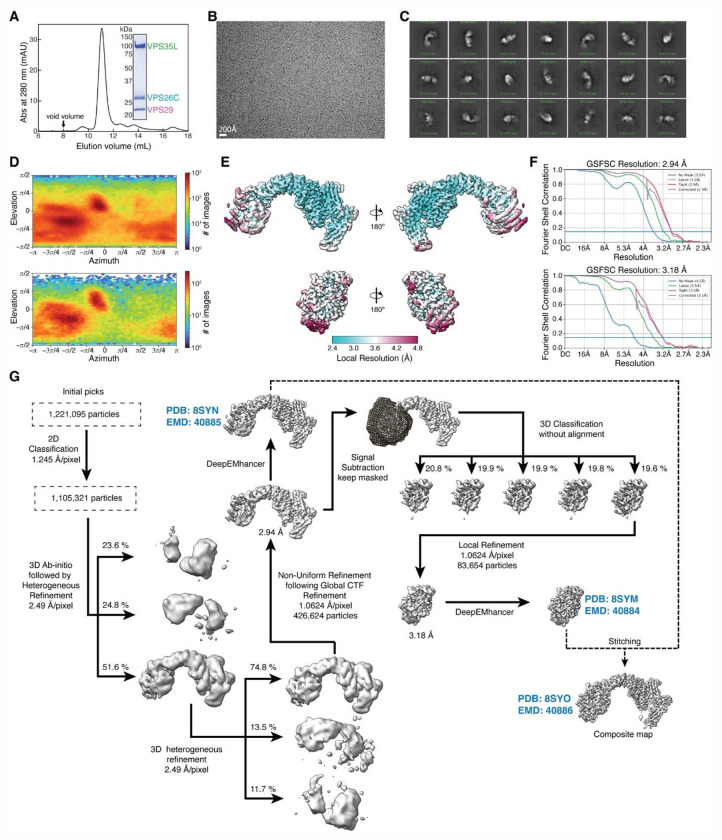
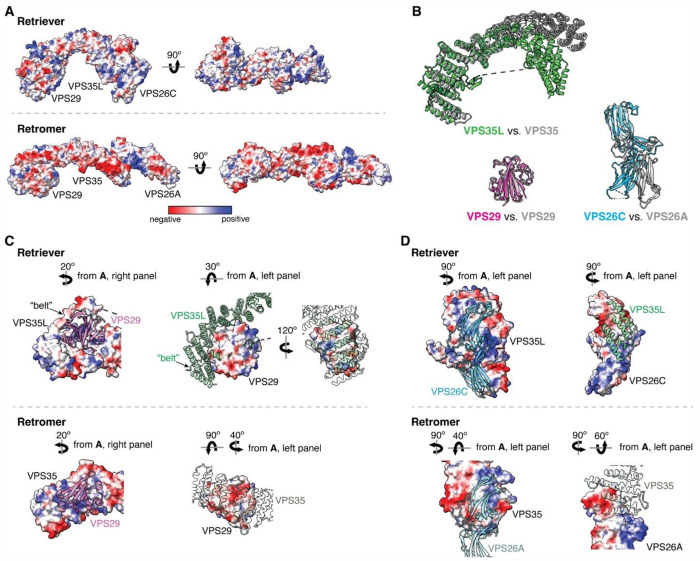
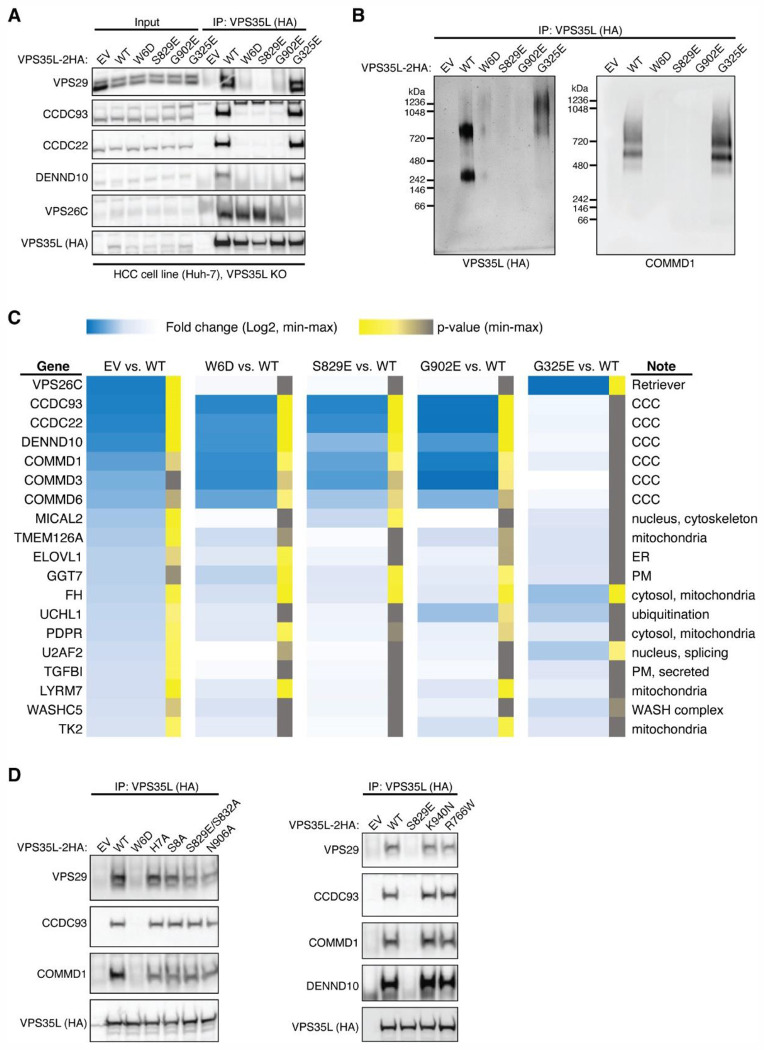
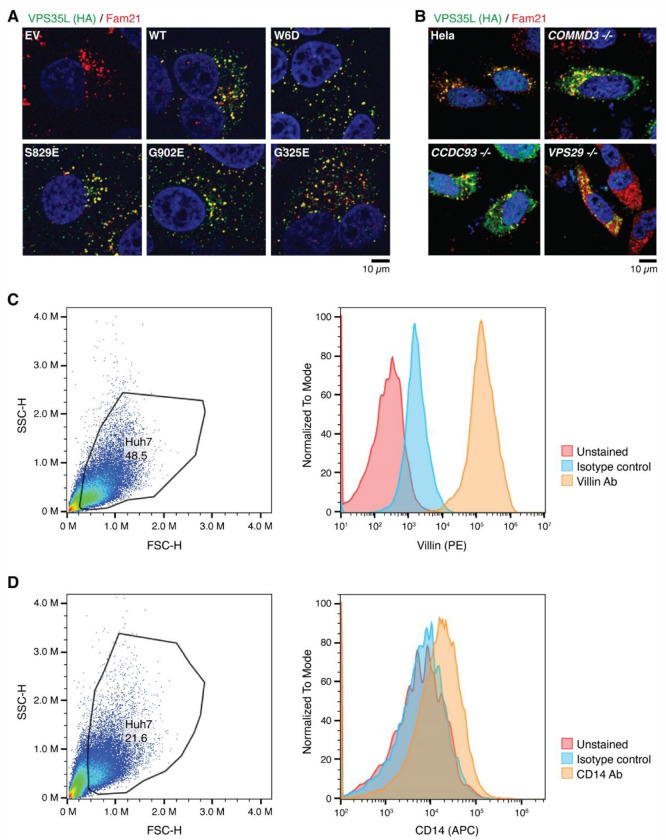
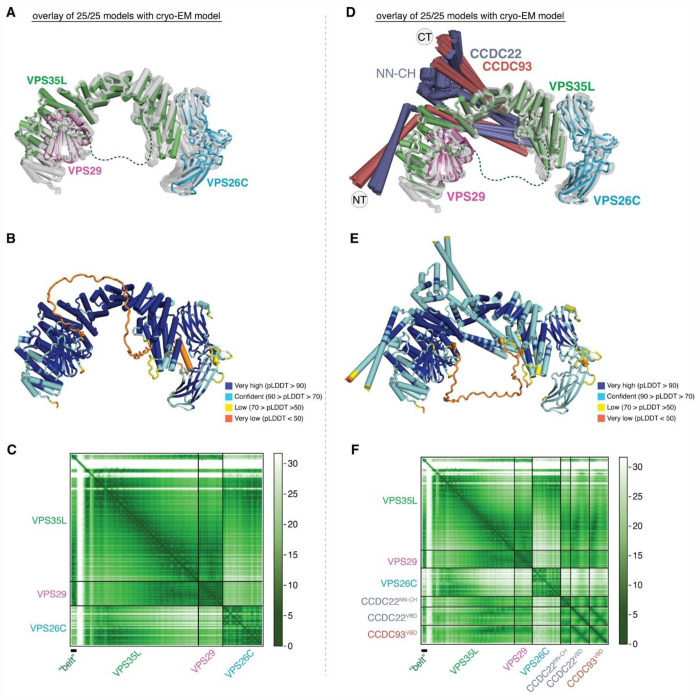
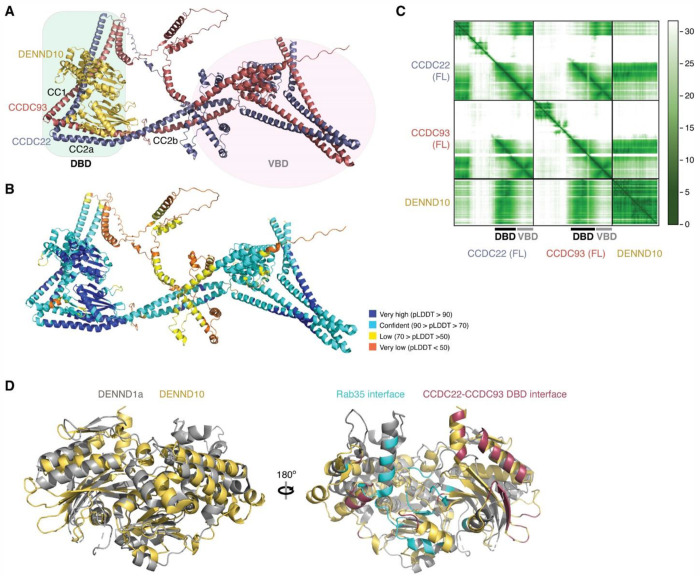
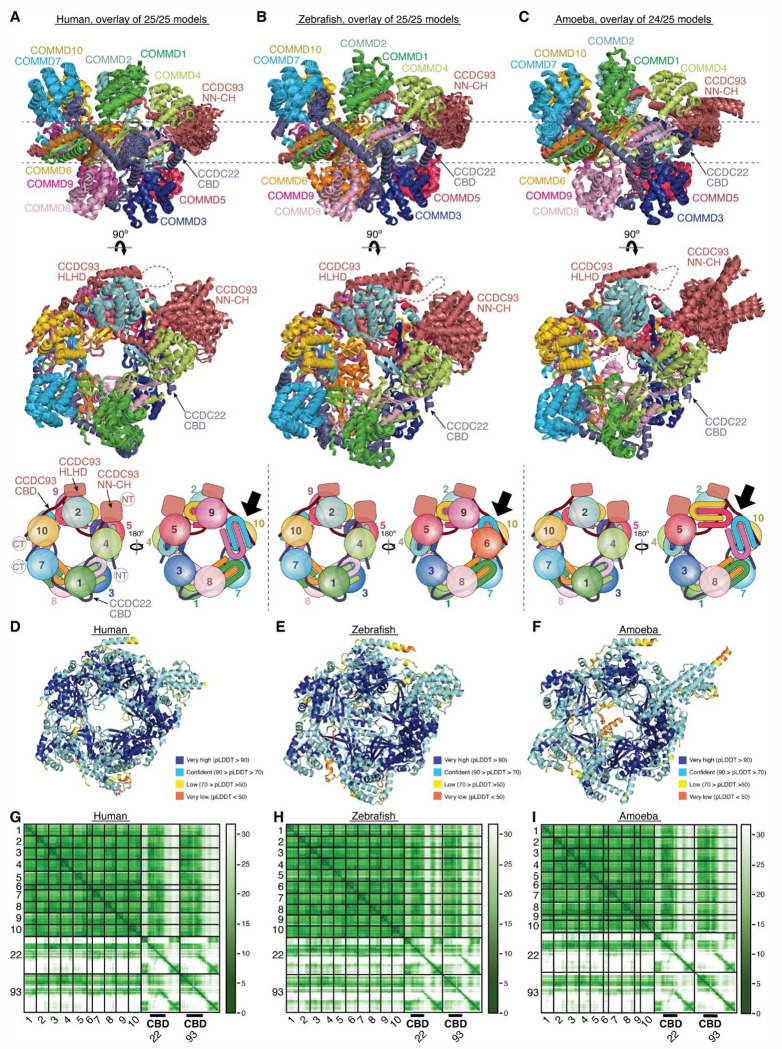
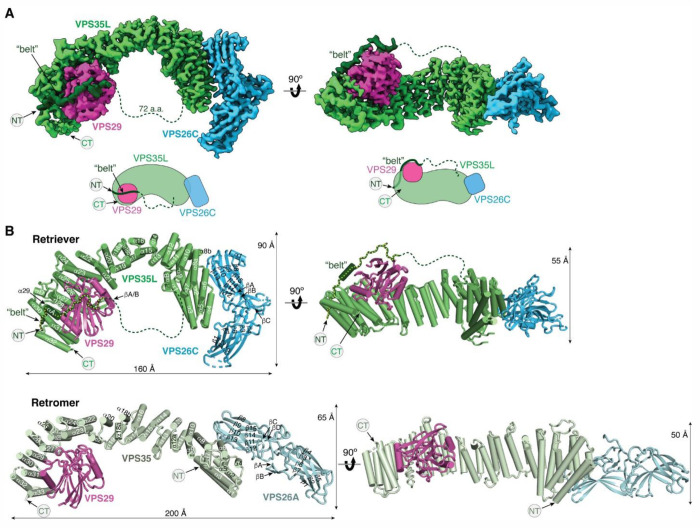
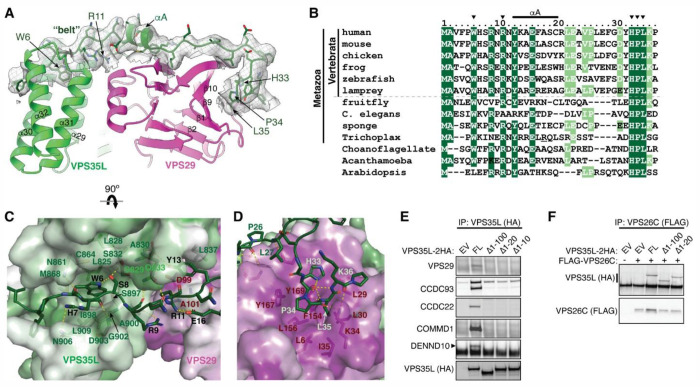
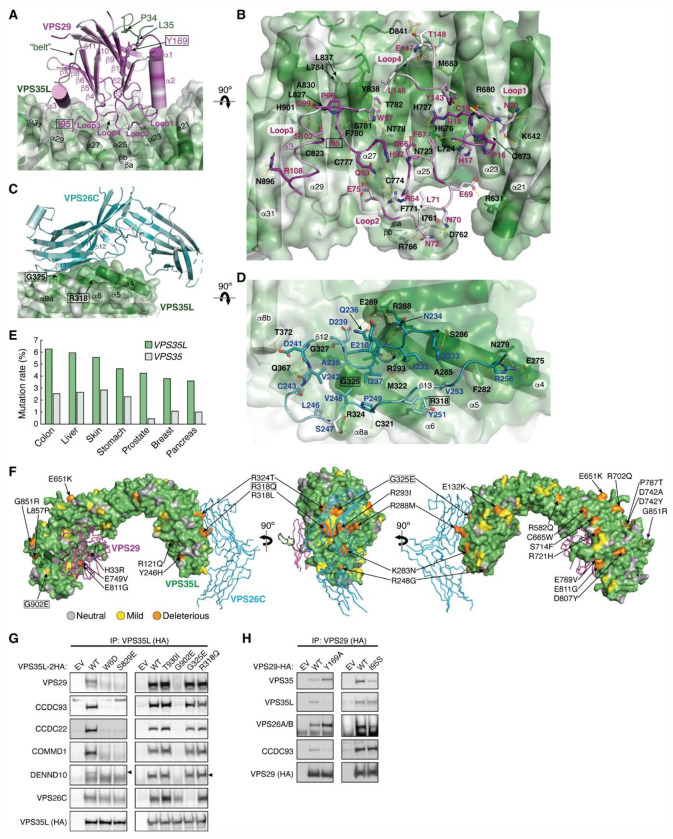
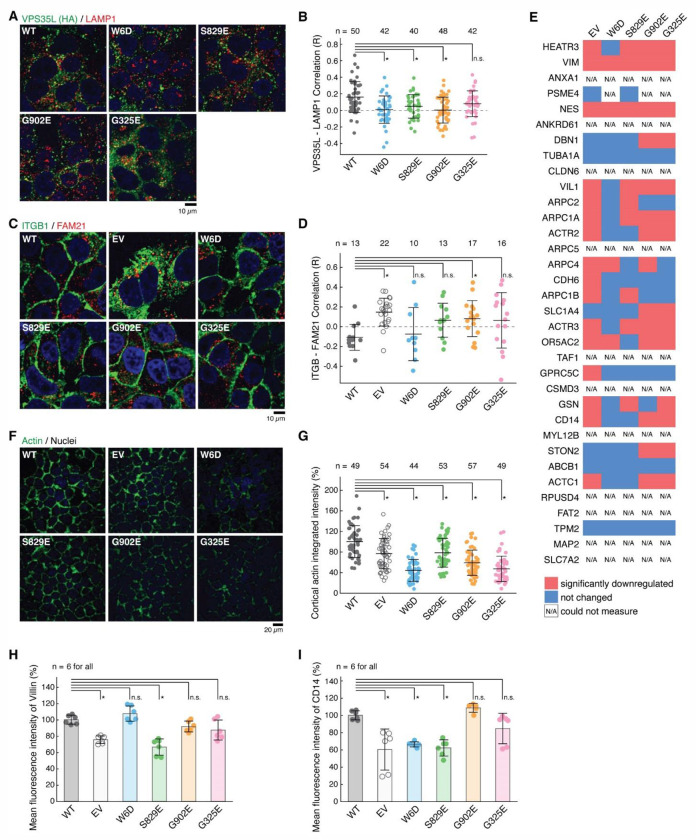
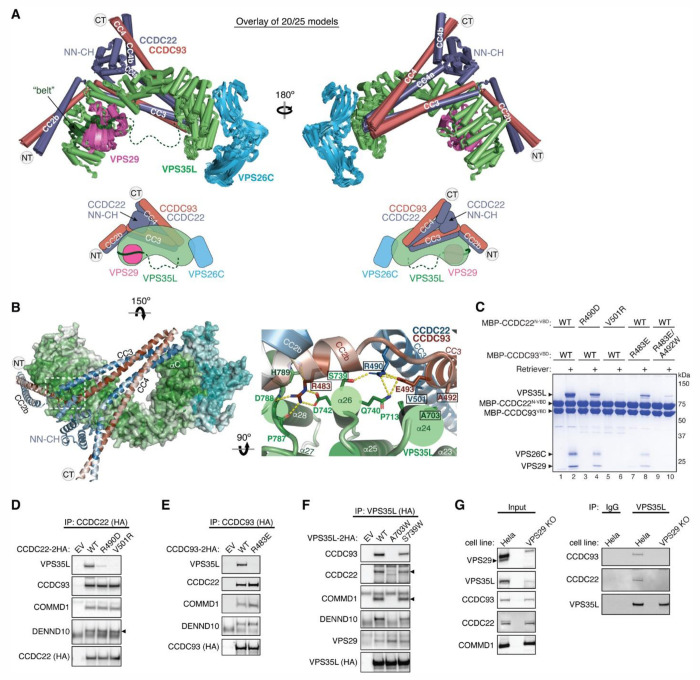
The recycling of membrane proteins from endosomes to the cell surface is vital for cell signaling and survival. Retriever, a trimeric complex of VPS35L, VPS26C and VPS29, together with the CCC complex comprising CCDC22, CCDC93, and COMMD proteins, plays a crucial role in this process. The precise mechanisms underlying Retriever assembly and its interaction with CCC have remained elusive. Here, we present the first high-resolution structure of Retriever determined using cryogenic electron microscopy. The structure reveals a unique assembly mechanism, distinguishing it from its remotely related paralog, Retromer. By combining AlphaFold predictions and biochemical, cellular, and proteomic analyses, we further elucidate the structural organization of the entire Retriever-CCC complex and uncover how cancer-associated mutations disrupt complex formation and impair membrane protein homeostasis. These findings provide a fundamental framework for understanding the biological and pathological implications associated with Retriever-CCC-mediated endosomal recycling.
Keywords: CCC complex; CCDC22; CCDC93; COMMD; Commander; Endosome recycling; Retriever; Retromer; VPS26C; VPS29; VPS35L.
Conflict of interest statement
Ethics Declarations The authors declare no competing interests.
Figures
Similar articles
-
Structural Organization of the Retriever-CCC Endosomal Recycling Complex.Res Sq [Preprint]. 2023 Jun 16:rs.3.rs-3026818. doi: 10.21203/rs.3.rs-3026818/v1. Res Sq. 2023. Update in: Nat Struct Mol Biol. 2024 Jun;31(6):910-924. doi: 10.1038/s41594-023-01184-4 PMID: 37397996 Free PMC article. Updated. Preprint.
-
Structural organization of the retriever-CCC endosomal recycling complex.Nat Struct Mol Biol. 2024 Jun;31(6):910-924. doi: 10.1038/s41594-023-01184-4. Epub 2023 Dec 7. Nat Struct Mol Biol. 2024. PMID: 38062209 Free PMC article.
-
Towards a molecular understanding of endosomal trafficking by Retromer and Retriever.Traffic. 2019 Jul;20(7):465-478. doi: 10.1111/tra.12649. Traffic. 2019. PMID: 30993794 Review.
-
Cargo-Specific Role for Retriever Subunit VPS26C in Hepatocyte Lipoprotein Receptor Recycling to Control Postprandial Triglyceride-Rich Lipoproteins.Arterioscler Thromb Vasc Biol. 2023 Jan;43(1):e29-e45. doi: 10.1161/ATVBAHA.122.318169. Epub 2022 Nov 10. Arterioscler Thromb Vasc Biol. 2023. PMID: 36353989
-
Commander Complex-A Multifaceted Operator in Intracellular Signaling and Cargo.Cells. 2021 Dec 7;10(12):3447. doi: 10.3390/cells10123447. Cells. 2021. PMID: 34943955 Free PMC article. Review.
References
-
- Uhlen M. et al. Proteomics. Tissue-based map of the human proteome. Science 347, 1260419 (2015). - PubMed
-
- Uhlen M. et al. Towards a knowledge-based Human Protein Atlas. Nat Biotechnol 28, 1248–50 (2010). - PubMed
-
- Edgar A.J. & Polak J.M. Human homologues of yeast vacuolar protein sorting 29 and 35. Biochem Biophys Res Commun 277, 622–30 (2000). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources