

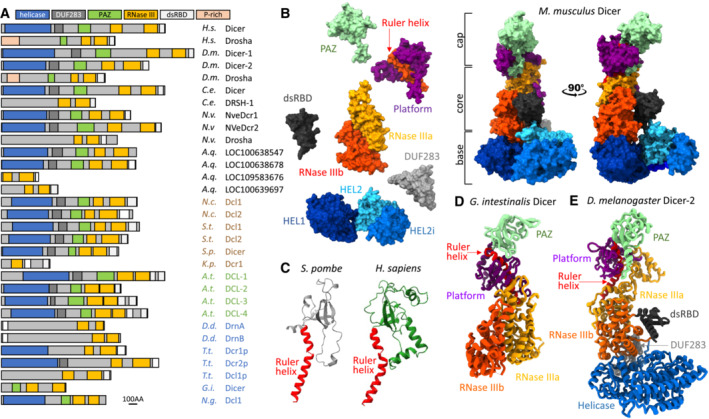
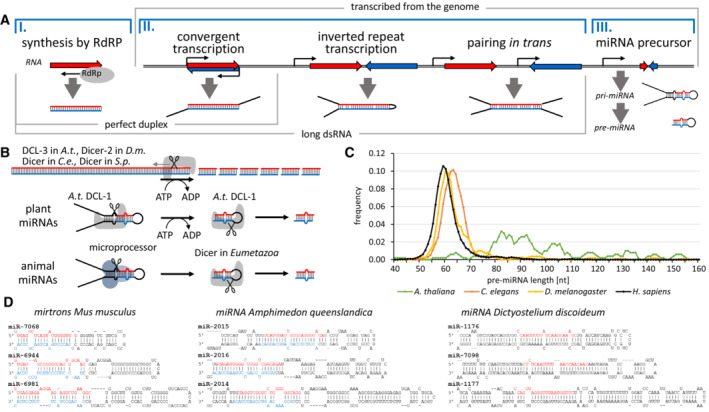
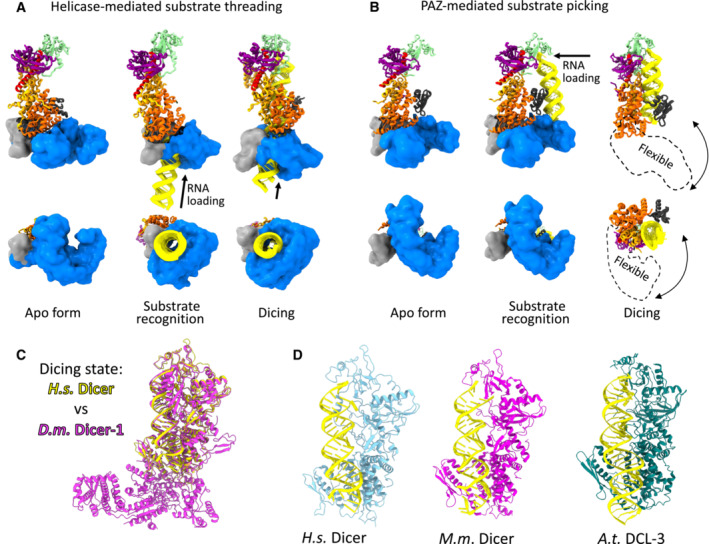
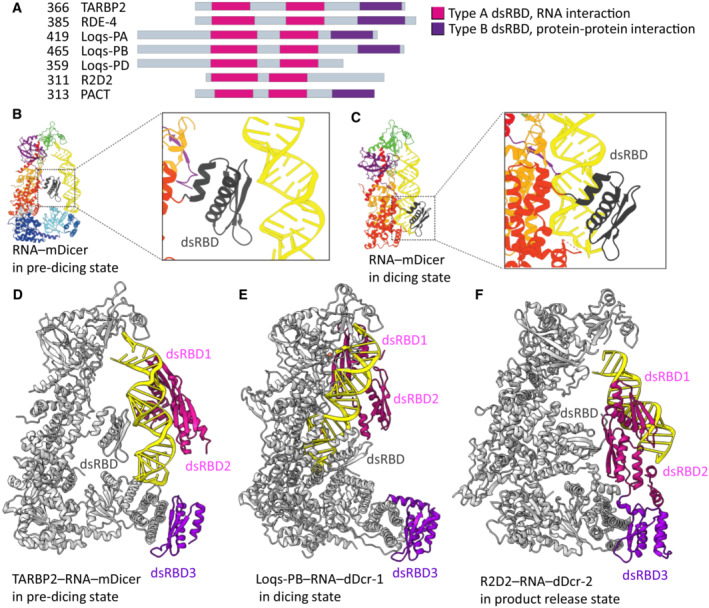
Dicer structure and function: conserved and evolving features
- PMID: 37310138
- PMCID: PMC10328071
- DOI: 10.15252/embr.202357215
Dicer structure and function: conserved and evolving features
Abstract
RNase III Dicer produces small RNAs guiding sequence-specific regulations, with important biological roles in eukaryotes. Major Dicer-dependent mechanisms are RNA interference (RNAi) and microRNA (miRNA) pathways, which employ distinct types of small RNAs. Small interfering RNAs (siRNAs) for RNAi are produced by Dicer from long double-stranded RNA (dsRNA) as a pool of different small RNAs. In contrast, miRNAs have specific sequences because they are precisely cleaved out from small hairpin precursors. Some Dicer homologs efficiently generate both, siRNAs and miRNAs, while others are adapted for biogenesis of one small RNA type. Here, we review the wealth of recent structural analyses of animal and plant Dicers, which have revealed how different domains and their adaptations contribute to substrate recognition and cleavage in different organisms and pathways. These data imply that siRNA generation was Dicer's ancestral role and that miRNA biogenesis relies on derived features. While the key element of functional divergence is a RIG-I-like helicase domain, Dicer-mediated small RNA biogenesis also documents the impressive functional versatility of the dsRNA-binding domain.
Keywords: Dicer; dsRBD; helicase; miRNA; siRNA.
© 2023 The Authors. Published under the terms of the CC BY 4.0 license.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Cryo-EM structures of human DICER dicing a pre-miRNA substrate.FEBS J. 2024 Jul;291(14):3072-3079. doi: 10.1111/febs.17048. Epub 2024 Jan 10. FEBS J. 2024. PMID: 38151772 Review.
-
Production of small RNAs by mammalian Dicer.Pflugers Arch. 2016 Jun;468(6):1089-102. doi: 10.1007/s00424-016-1817-6. Epub 2016 Apr 6. Pflugers Arch. 2016. PMID: 27048428 Free PMC article. Review.
-
The role of human Dicer-dsRBD in processing small regulatory RNAs.PLoS One. 2012;7(12):e51829. doi: 10.1371/journal.pone.0051829. Epub 2012 Dec 13. PLoS One. 2012. PMID: 23272173 Free PMC article.
-
Dicers at RISC; the mechanism of RNAi.Cell. 2004 Apr 2;117(1):1-3. doi: 10.1016/s0092-8674(04)00293-4. Cell. 2004. PMID: 15066275 Review.
-
Sequence determinant of small RNA production by DICER.Nature. 2023 Mar;615(7951):323-330. doi: 10.1038/s41586-023-05722-4. Epub 2023 Feb 22. Nature. 2023. PMID: 36813957
Cited by
-
Functional canonical RNAi in mice expressing a truncated Dicer isoform and long dsRNA.EMBO Rep. 2024 Jul;25(7):2896-2913. doi: 10.1038/s44319-024-00148-z. Epub 2024 May 20. EMBO Rep. 2024. PMID: 38769420 Free PMC article.
-
A Comprehensive Study of MicroRNA in Baculoviruses.Int J Mol Sci. 2024 Jan 3;25(1):603. doi: 10.3390/ijms25010603. Int J Mol Sci. 2024. PMID: 38203774 Free PMC article. Review.
-
Targeting Interleukin-17 as a Novel Treatment Option for Fibrotic Diseases.J Clin Med. 2023 Dec 27;13(1):164. doi: 10.3390/jcm13010164. J Clin Med. 2023. PMID: 38202170 Free PMC article. Review.
-
Antiviral Defence Mechanisms during Early Mammalian Development.Viruses. 2024 Jan 24;16(2):173. doi: 10.3390/v16020173. Viruses. 2024. PMID: 38399949 Free PMC article. Review.
-
Substrate promiscuity of Dicer toward precursors of the let-7 family and their 3'-end modifications.Cell Mol Life Sci. 2024 Jan 23;81(1):53. doi: 10.1007/s00018-023-05090-2. Cell Mol Life Sci. 2024. PMID: 38261114 Free PMC article.
References
-
- Bass BL, Hurst SR, Singer JD (1994) Binding properties of newly identified Xenopus proteins containing dsRNA‐binding motifs. Curr Biol 4: 301–314 - PubMed
-
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ (2001) Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 409: 363–366 - PubMed
-
- Blaszczyk J, Gan J, Tropea JE, Court DL, Waugh DS, Ji X (2004) Noncatalytic assembly of ribonuclease III with double‐stranded RNA. Structure 12: 457–466 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
