

The actin cytoskeleton in hair bundle development and hearing loss
- PMID: 37300948
- PMCID: PMC10408727
- DOI: 10.1016/j.heares.2023.108817
The actin cytoskeleton in hair bundle development and hearing loss
Abstract
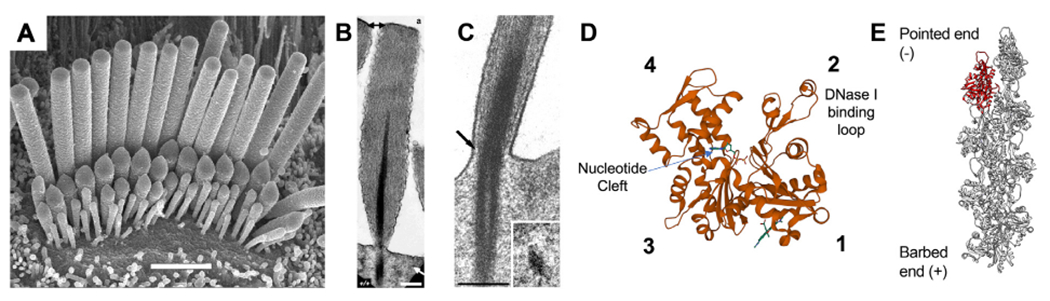
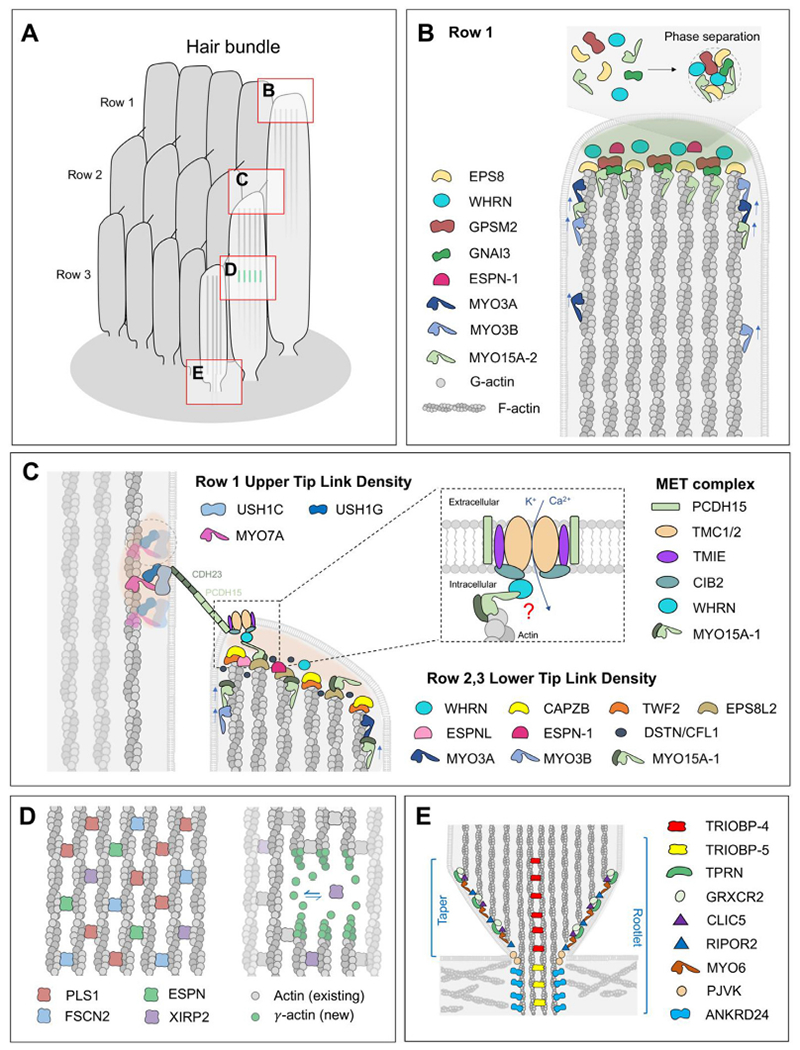
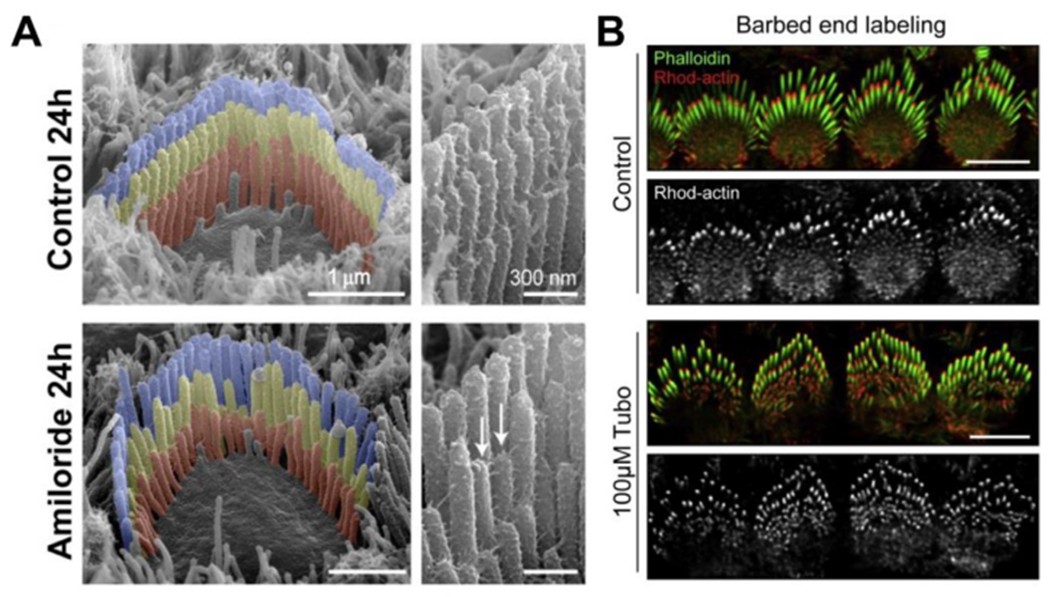
Inner ear hair cells assemble mechanosensitive hair bundles on their apical surface that transduce sounds and accelerations. Each hair bundle is comprised of ∼ 100 individual stereocilia that are arranged into rows of increasing height and width; their specific and precise architecture being necessary for mechanoelectrical transduction (MET). The actin cytoskeleton is fundamental to establishing this architecture, not only by forming the structural scaffold shaping each stereocilium, but also by composing rootlets and the cuticular plate that together provide a stable foundation supporting each stereocilium. In concert with the actin cytoskeleton, a large assortment of actin-binding proteins (ABPs) function to cross-link actin filaments into specific topologies, as well as control actin filament growth, severing, and capping. These processes are individually critical for sensory transduction and are all disrupted in hereditary forms of human hearing loss. In this review, we provide an overview of actin-based structures in the hair bundle and the molecules contributing to their assembly and functional properties. We also highlight recent advances in mechanisms driving stereocilia elongation and how these processes are tuned by MET.
Keywords: Actin; Actin binding protein; Deafness; Hair bundle; Hearing loss; Stereocilia.
Copyright © 2023 Elsevier B.V. All rights reserved.
Figures
Similar articles
-
Correlation of expression of the actin filament-bundling protein espin with stereociliary bundle formation in the developing inner ear.J Comp Neurol. 2004 Jan 1;468(1):125-34. doi: 10.1002/cne.10944. J Comp Neurol. 2004. PMID: 14648695
-
The deaf mouse mutant whirler suggests a role for whirlin in actin filament dynamics and stereocilia development.Cell Motil Cytoskeleton. 2007 Jul;64(7):496-508. doi: 10.1002/cm.20199. Cell Motil Cytoskeleton. 2007. PMID: 17326148 Free PMC article.
-
Actin-bundling protein TRIOBP forms resilient rootlets of hair cell stereocilia essential for hearing.Cell. 2010 May 28;141(5):786-98. doi: 10.1016/j.cell.2010.03.049. Cell. 2010. PMID: 20510926 Free PMC article.
-
Stereocilia Rootlets: Actin-Based Structures That Are Essential for Structural Stability of the Hair Bundle.Int J Mol Sci. 2020 Jan 3;21(1):324. doi: 10.3390/ijms21010324. Int J Mol Sci. 2020. PMID: 31947734 Free PMC article. Review.
-
The cuticular plate: a riddle, wrapped in a mystery, inside a hair cell.Birth Defects Res C Embryo Today. 2015 Jun;105(2):126-39. doi: 10.1002/bdrc.21098. Epub 2015 Jun 23. Birth Defects Res C Embryo Today. 2015. PMID: 26104653 Review.
Cited by
-
What are the main proteins in the hemolymph of Haemaphysalis flava ticks?Front Vet Sci. 2024 Jul 17;11:1387719. doi: 10.3389/fvets.2024.1387719. eCollection 2024. Front Vet Sci. 2024. PMID: 39086760 Free PMC article.
-
The Relevance of Autophagy within Inner Ear in Baseline Conditions and Tinnitus-Related Syndromes.Int J Mol Sci. 2023 Nov 23;24(23):16664. doi: 10.3390/ijms242316664. Int J Mol Sci. 2023. PMID: 38068993 Free PMC article. Review.
-
Organization of a cytoskeletal superstructure in the apical domain of intestinal tuft cells.J Cell Biol. 2024 Dec 2;223(12):e202404070. doi: 10.1083/jcb.202404070. Epub 2024 Oct 1. J Cell Biol. 2024. PMID: 39352498 Free PMC article.
-
Keratan sulfate, an electrosensory neurosentient bioresponsive cell instructive glycosaminoglycan.Glycobiology. 2024 Apr 1;34(3):cwae014. doi: 10.1093/glycob/cwae014. Glycobiology. 2024. PMID: 38376199 Free PMC article. Review.
References
-
- Abu Rayyan A, Kamal L, Casadei S, Brownstein Z, Zahdeh F, Shahin H, Canavati C, Dweik D, Jaraysa T, Rabie G, Carlson RJ, Gulsuner S, Lee MK, Avraham KB, Walsh T, King M-C, Kanaan MN, 2020. Genomic analysis of inherited hearing loss in the Palestinian population. Proc. Natl. Acad. Sci. U.S.A 117, 20070–20076. doi:10.1073/pnas.2009628117. - DOI - PMC - PubMed
-
- Avenarius MR, Krey JF, Dumont RA, Morgan CP, Benson CB, Vijayakumar S, Cunningham CL, Scheffer DI, Corey DP, Müller U, Jones SM, Barr-Gillespie PG, 2017. Heterodimeric capping protein is required for stereocilia length and width regulation. J. Cell Biol 216, 3861–3881. doi:10.1083/jcb.201704171. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
