

Therapeutic targeting of metabolic vulnerabilities in cancers with MLL3/4-COMPASS epigenetic regulator mutations
- PMID: 37252797
- PMCID: PMC10313365
- DOI: 10.1172/JCI169993
Therapeutic targeting of metabolic vulnerabilities in cancers with MLL3/4-COMPASS epigenetic regulator mutations
Abstract
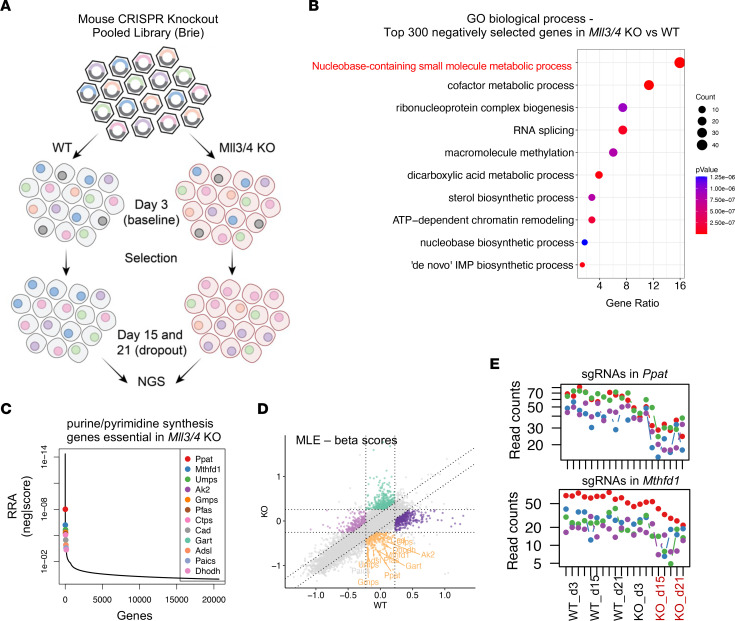
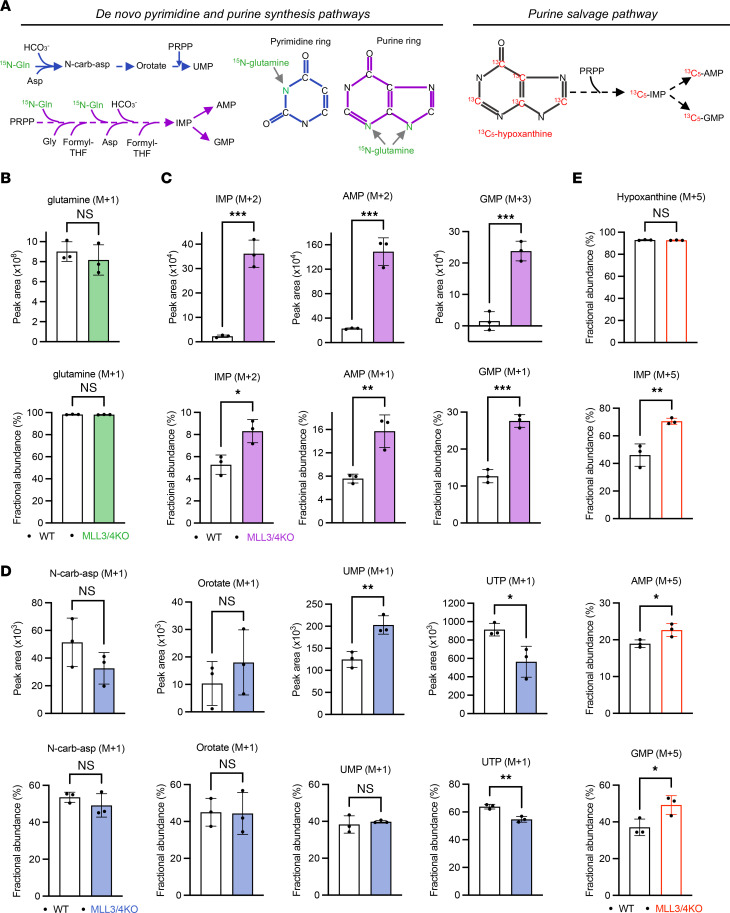
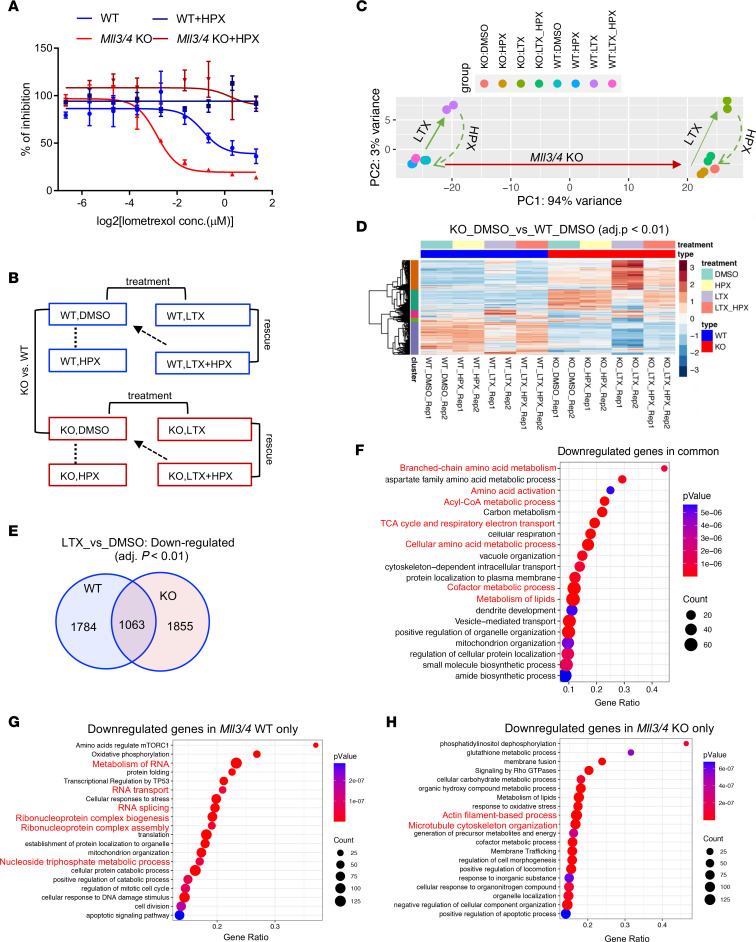
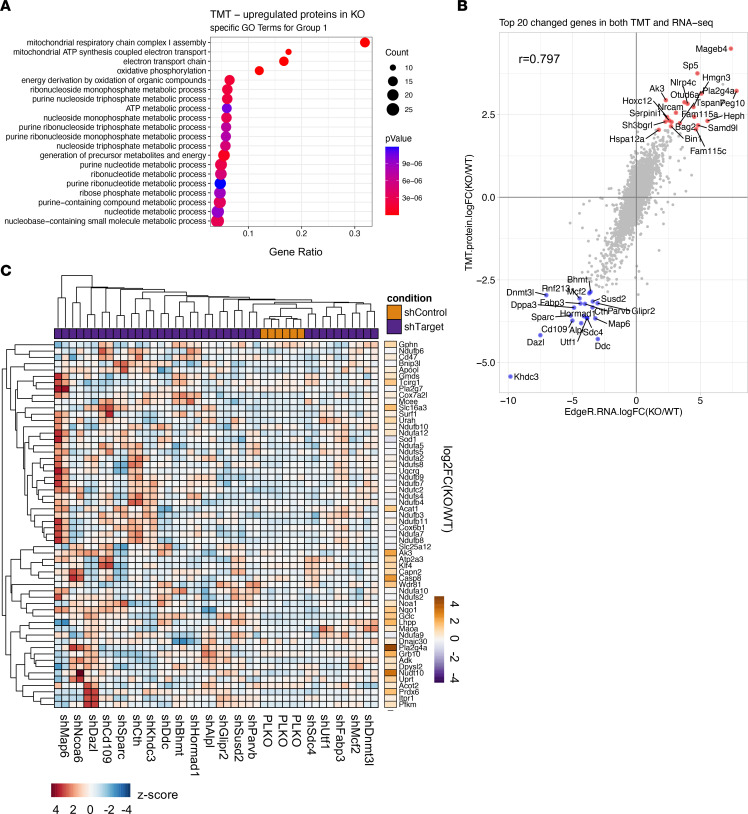
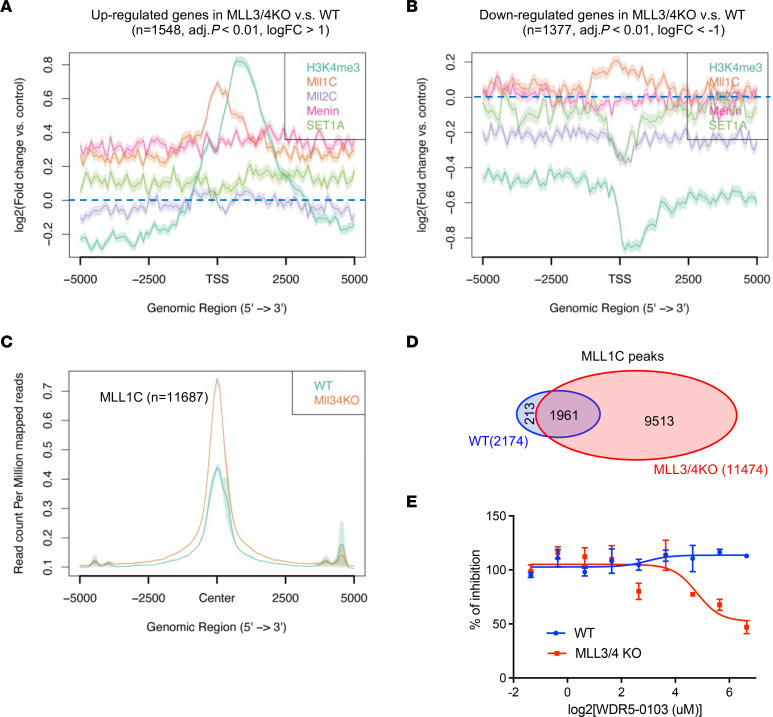
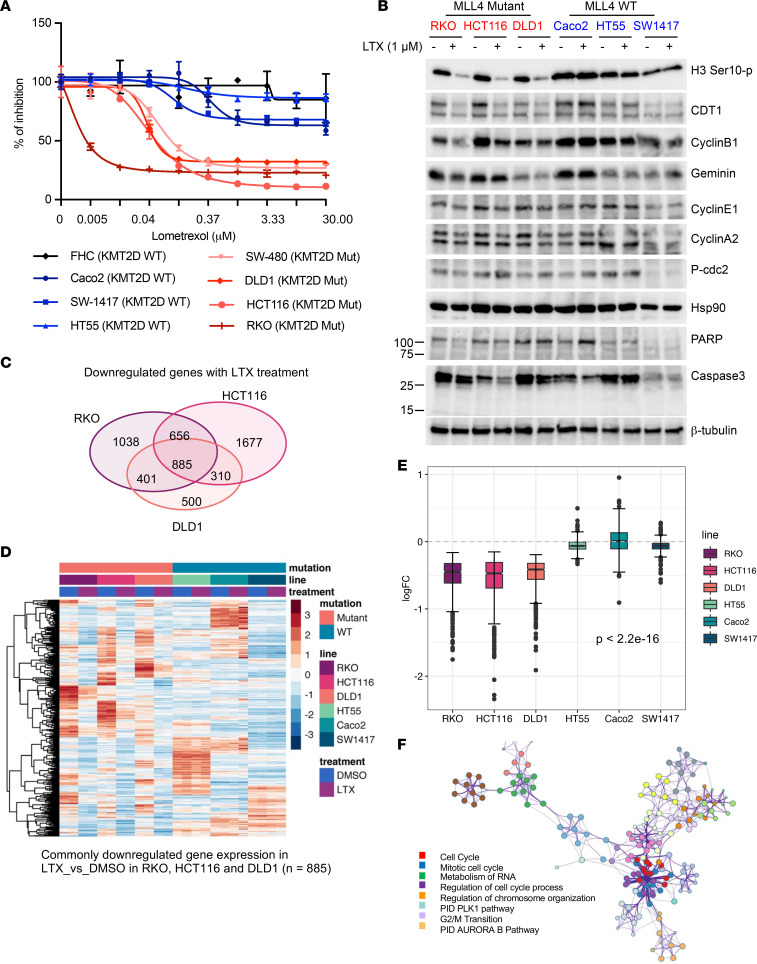
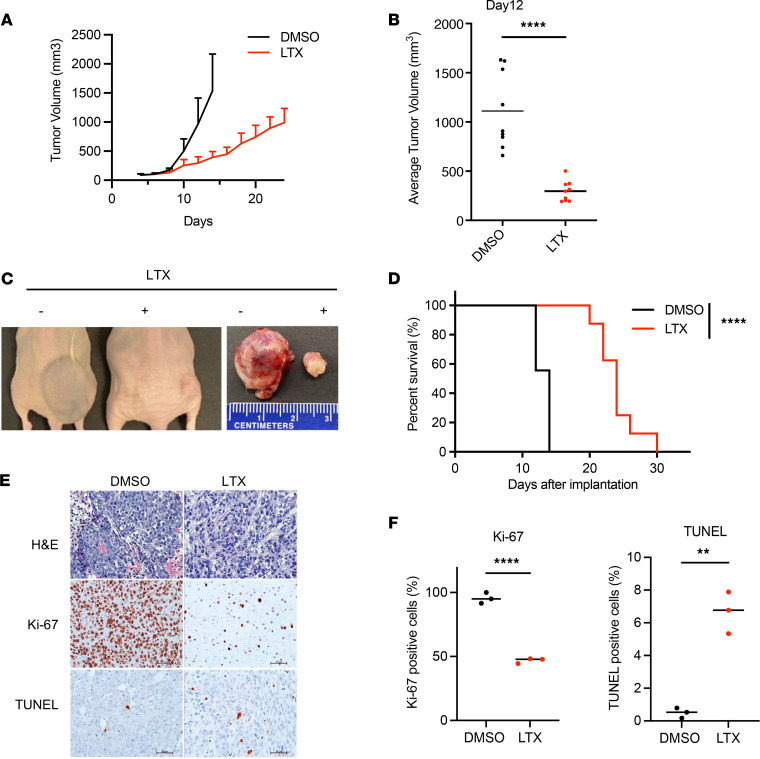
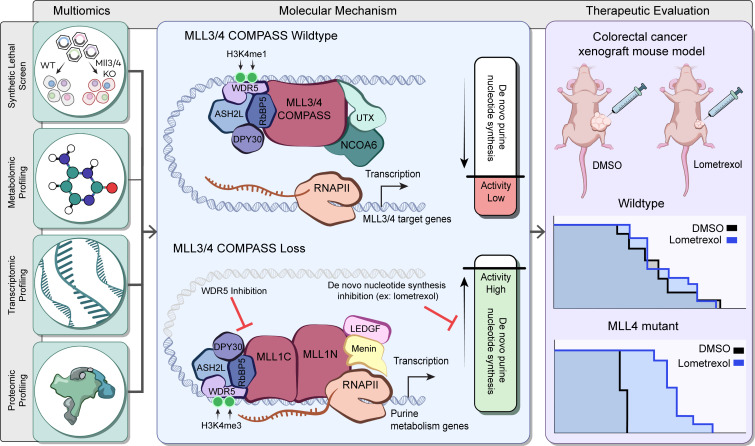
Epigenetic status-altering mutations in chromatin-modifying enzymes are a feature of human diseases, including many cancers. However, the functional outcomes and cellular dependencies arising from these mutations remain unresolved. In this study, we investigated cellular dependencies, or vulnerabilities, that arise when enhancer function is compromised by loss of the frequently mutated COMPASS family members MLL3 and MLL4. CRISPR dropout screens in MLL3/4-depleted mouse embryonic stem cells (mESCs) revealed synthetic lethality upon suppression of purine and pyrimidine nucleotide synthesis pathways. Consistently, we observed a shift in metabolic activity toward increased purine synthesis in MLL3/4-KO mESCs. These cells also exhibited enhanced sensitivity to the purine synthesis inhibitor lometrexol, which induced a unique gene expression signature. RNA-Seq identified the top MLL3/4 target genes coinciding with suppression of purine metabolism, and tandem mass tag proteomic profiling further confirmed upregulation of purine synthesis in MLL3/4-KO cells. Mechanistically, we demonstrated that compensation by MLL1/COMPASS was underlying these effects. Finally, we demonstrated that tumors with MLL3 and/or MLL4 mutations were highly sensitive to lometrexol in vitro and in vivo, both in culture and in animal models of cancer. Our results depicted a targetable metabolic dependency arising from epigenetic factor deficiency, providing molecular insight to inform therapy for cancers with epigenetic alterations secondary to MLL3/4 COMPASS dysfunction.
Keywords: Colorectal cancer; Epigenetics; Genetics; Metabolism.
Conflict of interest statement
Figures
Similar articles
-
The MLL3/MLL4 branches of the COMPASS family function as major histone H3K4 monomethylases at enhancers.Mol Cell Biol. 2013 Dec;33(23):4745-54. doi: 10.1128/MCB.01181-13. Epub 2013 Sep 30. Mol Cell Biol. 2013. PMID: 24081332 Free PMC article.
-
The MLL3/4 H3K4 methyltransferase complex in establishing an active enhancer landscape.Biochem Soc Trans. 2021 Jun 30;49(3):1041-1054. doi: 10.1042/BST20191164. Biochem Soc Trans. 2021. PMID: 34156443 Free PMC article. Review.
-
A small UTX stabilization domain of Trr is conserved within mammalian MLL3-4/COMPASS and is sufficient to rescue loss of viability in null animals.Genes Dev. 2020 Nov 1;34(21-22):1493-1502. doi: 10.1101/gad.339762.120. Epub 2020 Oct 8. Genes Dev. 2020. PMID: 33033055 Free PMC article.
-
Resetting the epigenetic balance of Polycomb and COMPASS function at enhancers for cancer therapy.Nat Med. 2018 Jun;24(6):758-769. doi: 10.1038/s41591-018-0034-6. Epub 2018 May 21. Nat Med. 2018. PMID: 29785026 Free PMC article.
-
COMPASS Ascending: Emerging clues regarding the roles of MLL3/KMT2C and MLL2/KMT2D proteins in cancer.Cancer Lett. 2019 Aug 28;458:56-65. doi: 10.1016/j.canlet.2019.05.024. Epub 2019 May 22. Cancer Lett. 2019. PMID: 31128216 Free PMC article. Review.
Cited by
-
Epigenetic therapies targeting histone lysine methylation: complex mechanisms and clinical challenges.J Clin Invest. 2024 Oct 15;134(20):e183391. doi: 10.1172/JCI183391. J Clin Invest. 2024. PMID: 39403928 Free PMC article. Review.
-
Somatic mutations of MLL4/COMPASS induce cytoplasmic localization providing molecular insight into cancer prognosis and treatment.Proc Natl Acad Sci U S A. 2023 Dec 26;120(52):e2310063120. doi: 10.1073/pnas.2310063120. Epub 2023 Dec 19. Proc Natl Acad Sci U S A. 2023. PMID: 38113256 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
