

An Endogenous Retrovirus Vaccine Encoding an Envelope with a Mutated Immunosuppressive Domain in Combination with Anti-PD1 Treatment Eradicates Established Tumours in Mice
- PMID: 37112906
- PMCID: PMC10141008
- DOI: 10.3390/v15040926
An Endogenous Retrovirus Vaccine Encoding an Envelope with a Mutated Immunosuppressive Domain in Combination with Anti-PD1 Treatment Eradicates Established Tumours in Mice
Abstract
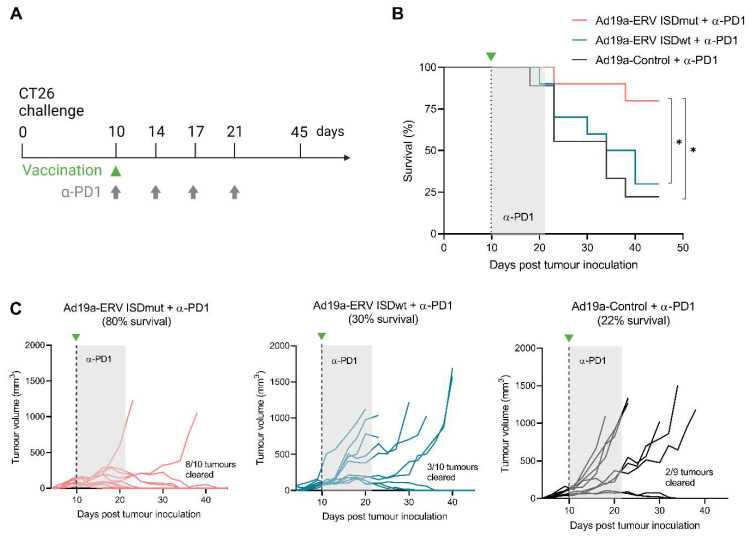
Endogenous retroviruses (ERVs) account for 8% of our genome, and, although they are usually silent in healthy tissues, they become reactivated and expressed in pathological conditions such as cancer. Several studies support a functional role of ERVs in tumour development and progression, specifically through their envelope (Env) protein, which contains a region described as an immunosuppressive domain (ISD). We have previously shown that targeting of the murine ERV (MelARV) Env using virus-like vaccine (VLV) technology, consisting of an adenoviral vector encoding virus-like particles (VLPs), induces protection against small tumours in mice. Here, we investigate the potency and efficacy of a novel MelARV VLV with a mutated ISD (ISDmut) that can modify the properties of the adenoviral vaccine-encoded Env protein. We show that the modification of the vaccine's ISD significantly enhanced T-cell immunogenicity in both prime and prime-boost vaccination regimens. The modified VLV in combination with an α-PD1 checkpoint inhibitor (CPI) exhibited excellent curative efficacy against large established colorectal CT26 tumours in mice. Furthermore, only ISDmut-vaccinated mice that survived CT26 challenge were additionally protected against rechallenge with a triple-negative breast cancer cell line (4T1), showing that our modified VLV provides cross-protection against different tumour types expressing ERV-derived antigens. We envision that translating these findings and technology into human ERVs (HERVs) could provide new treatment opportunities for cancer patients with unmet medical needs.
Keywords: adenoviral vectors; cancer; endogenous retroviruses; immunotherapy; murine melanoma-associated retrovirus; virus-like particles; virus-like-vaccines.
Conflict of interest statement
The project was sponsored by InProTher ApS, in collaboration with Sirion Biotech, that provided the used vaccines. L.N., P.J.H. and C.T. are co-inventors of the virus-like vaccine technology for immunotherapy against cancer. P.J.H. is the current CSO of InProTher ApS and has had the company as his primary employer since January 2019. P.J.H. and C.T. are major shareholders and board members of InProTher ApS. J.D., E.R., I.S., K.N.N., A.V.B., A.A. and L.N have been or are current employees and warrant holders of InProTher ApS. Despite this, the aforementioned facts had no influence on the design of the experimental experiments, nor the data and their representation.
Figures


Similar articles
-
Human Ad19a/64 HERV-W Vaccines Uncover Immunosuppression Domain-Dependent T-Cell Response Differences in Inbred Mice.Int J Mol Sci. 2023 Jun 9;24(12):9972. doi: 10.3390/ijms24129972. Int J Mol Sci. 2023. PMID: 37373123 Free PMC article.
-
Adenovirus based virus-like-vaccines targeting endogenous retroviruses can eliminate growing colorectal cancers in mice.Oncotarget. 2019 Feb 15;10(14):1458-1472. doi: 10.18632/oncotarget.26680. eCollection 2019 Feb 15. Oncotarget. 2019. PMID: 30858929 Free PMC article.
-
Cancer Associated Endogenous Retroviruses: Ideal Immune Targets for Adenovirus-Based Immunotherapy.Int J Mol Sci. 2020 Jul 8;21(14):4843. doi: 10.3390/ijms21144843. Int J Mol Sci. 2020. PMID: 32650622 Free PMC article. Review.
-
Human endogenous retroviruses and cancer prevention: evidence and prospects.BMC Cancer. 2013 Jan 3;13:4. doi: 10.1186/1471-2407-13-4. BMC Cancer. 2013. PMID: 23282240 Free PMC article.
-
Murine polyomavirus virus-like particles (VLPs) as vectors for gene and immune therapy and vaccines against viral infections and cancer.Anticancer Res. 2005 Jul-Aug;25(4):2601-8. Anticancer Res. 2005. PMID: 16080500 Review.
Cited by
-
Exploring HERV-K (HML-2) Influence in Cancer and Prospects for Therapeutic Interventions.Int J Mol Sci. 2023 Sep 27;24(19):14631. doi: 10.3390/ijms241914631. Int J Mol Sci. 2023. PMID: 37834078 Free PMC article. Review.
-
The Insertion of an Evolutionary Lost Four-Amino-Acid Cytoplasmic Tail Peptide into a Syncytin-1 Vaccine Increases T- and B-Cell Responses in Mice.Viruses. 2023 Aug 3;15(8):1686. doi: 10.3390/v15081686. Viruses. 2023. PMID: 37632028 Free PMC article.
-
Human Ad19a/64 HERV-W Vaccines Uncover Immunosuppression Domain-Dependent T-Cell Response Differences in Inbred Mice.Int J Mol Sci. 2023 Jun 9;24(12):9972. doi: 10.3390/ijms24129972. Int J Mol Sci. 2023. PMID: 37373123 Free PMC article.
-
The Role of Human Endogenous Retroviruses in Cancer Immunotherapy of the Post-COVID-19 World.Cancers (Basel). 2023 Nov 7;15(22):5321. doi: 10.3390/cancers15225321. Cancers (Basel). 2023. PMID: 38001581 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
