

Polyunsaturated fatty acids-rich dietary lipid prevents high fat diet-induced obesity in mice
- PMID: 37019935
- PMCID: PMC10076282
- DOI: 10.1038/s41598-023-32851-7
Polyunsaturated fatty acids-rich dietary lipid prevents high fat diet-induced obesity in mice
Abstract
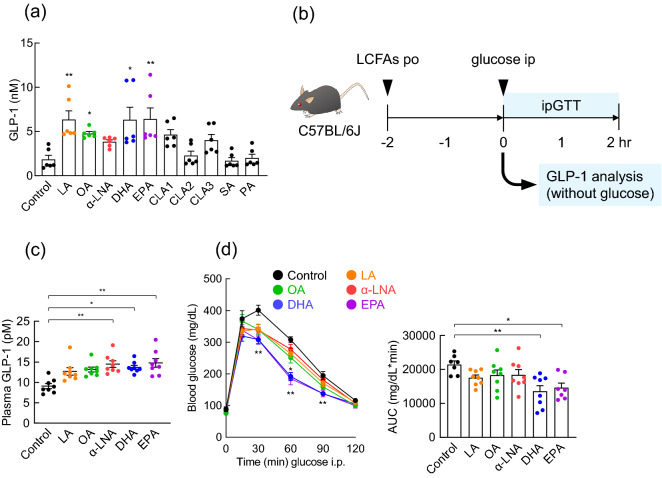
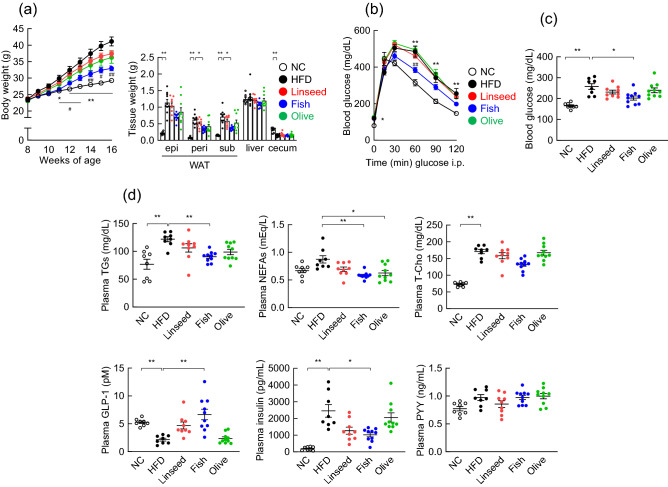
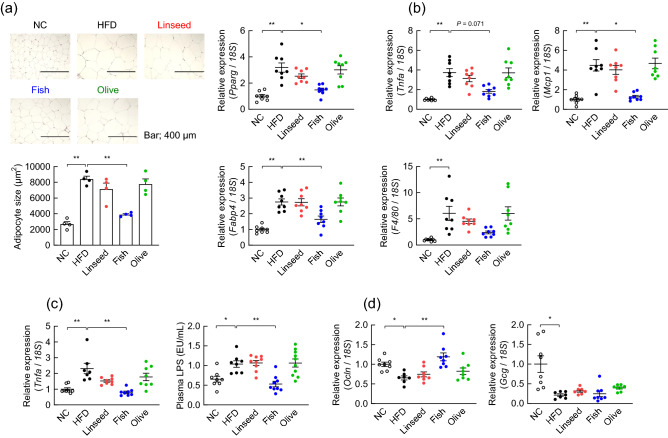
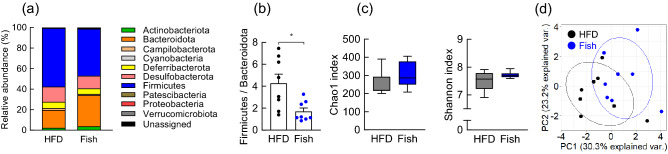
Diet is the primary factor affecting host nutrition and metabolism, with excess food intake, especially high-calorie diets, such as high-fat and high-sugar diets, causing an increased risk of obesity and related disorders. Obesity alters the gut microbial composition and reduces microbial diversity and causes changes in specific bacterial taxa. Dietary lipids can alter the gut microbial composition in obese mice. However, the regulation of gut microbiota and host energy homeostasis by different polyunsaturated fatty acids (PUFAs) in dietary lipids remains unknown. Here, we demonstrated that different PUFAs in dietary lipids improved host metabolism in high-fat diet (HFD)-induced obesity in mice. The intake of the different PUFA-enriched dietary lipids improved metabolism in HFD-induced obesity by regulating glucose tolerance and inhibiting colonic inflammation. Moreover, the gut microbial compositions were different among HFD and modified PUFA-enriched HFD-fed mice. Thus, we have identified a new mechanism underlying the function of different PUFAs in dietary lipids in regulating host energy homeostasis in obese conditions. Our findings shed light on the prevention and treatment of metabolic disorders by targeting the gut microbiota.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Gut microbiota confers host resistance to obesity by metabolizing dietary polyunsaturated fatty acids.Nat Commun. 2019 Sep 5;10(1):4007. doi: 10.1038/s41467-019-11978-0. Nat Commun. 2019. PMID: 31488836 Free PMC article.
-
Fat and not sugar as the determining factor for gut microbiota changes, obesity, and related metabolic disorders in mice.Am J Physiol Endocrinol Metab. 2023 Jan 1;324(1):E85-E96. doi: 10.1152/ajpendo.00141.2022. Epub 2022 Dec 14. Am J Physiol Endocrinol Metab. 2023. PMID: 36516223
-
Composition of dietary fat source shapes gut microbiota architecture and alters host inflammatory mediators in mouse adipose tissue.JPEN J Parenter Enteral Nutr. 2013 Nov;37(6):746-54. doi: 10.1177/0148607113486931. Epub 2013 May 2. JPEN J Parenter Enteral Nutr. 2013. PMID: 23639897 Free PMC article.
-
Dietary Fat and Cancer-Which Is Good, Which Is Bad, and the Body of Evidence.Int J Mol Sci. 2020 Jun 9;21(11):4114. doi: 10.3390/ijms21114114. Int J Mol Sci. 2020. PMID: 32526973 Free PMC article. Review.
-
Effect of dietary fatty acid composition on substrate utilization and body weight maintenance in humans.Eur J Nutr. 2014 Apr;53(3):691-710. doi: 10.1007/s00394-013-0638-z. Epub 2013 Dec 22. Eur J Nutr. 2014. PMID: 24363161 Review.
Cited by
-
Brain Perception of Different Oils on Appetite Regulation: An Anorectic Gene Expression Pattern in the Hypothalamus Dependent on the Vagus Nerve.Nutrients. 2024 Jul 24;16(15):2397. doi: 10.3390/nu16152397. Nutrients. 2024. PMID: 39125278 Free PMC article.
-
The Role of Organokines in Obesity and Type 2 Diabetes and Their Functions as Molecular Transducers of Nutrition and Exercise.Metabolites. 2023 Aug 29;13(9):979. doi: 10.3390/metabo13090979. Metabolites. 2023. PMID: 37755259 Free PMC article. Review.
-
Recent insights of obesity-induced gut and adipose tissue dysbiosis in type 2 diabetes.Front Mol Biosci. 2023 Sep 28;10:1224982. doi: 10.3389/fmolb.2023.1224982. eCollection 2023. Front Mol Biosci. 2023. PMID: 37842639 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
