

Neuronal activity regulates Matrin 3 abundance and function in a calcium-dependent manner through calpain-mediated cleavage and calmodulin binding
- PMID: 37011198
- PMCID: PMC10104577
- DOI: 10.1073/pnas.2206217120
Neuronal activity regulates Matrin 3 abundance and function in a calcium-dependent manner through calpain-mediated cleavage and calmodulin binding
Abstract
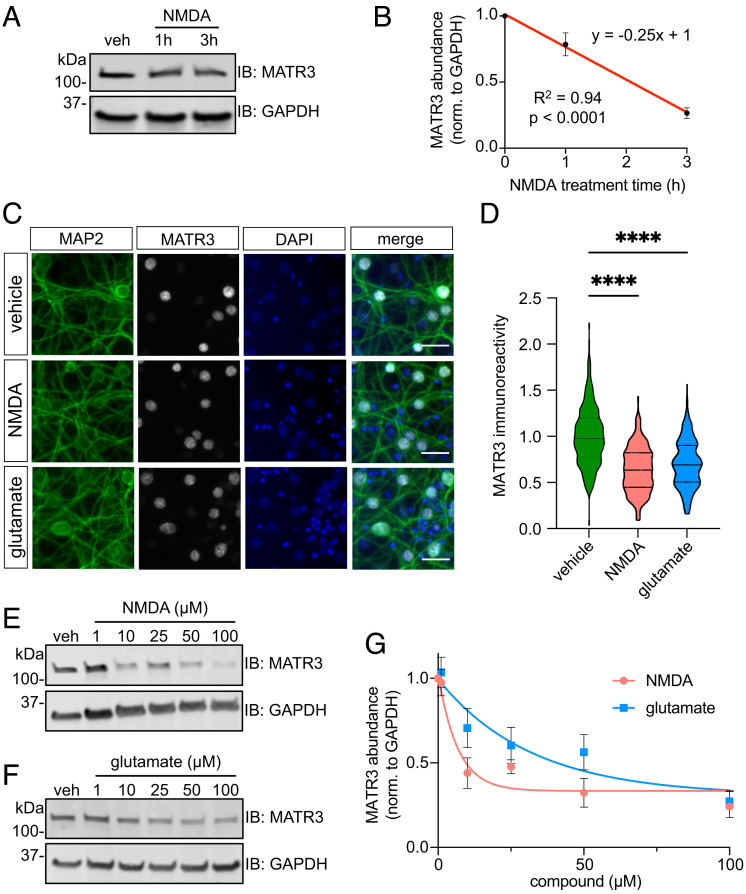
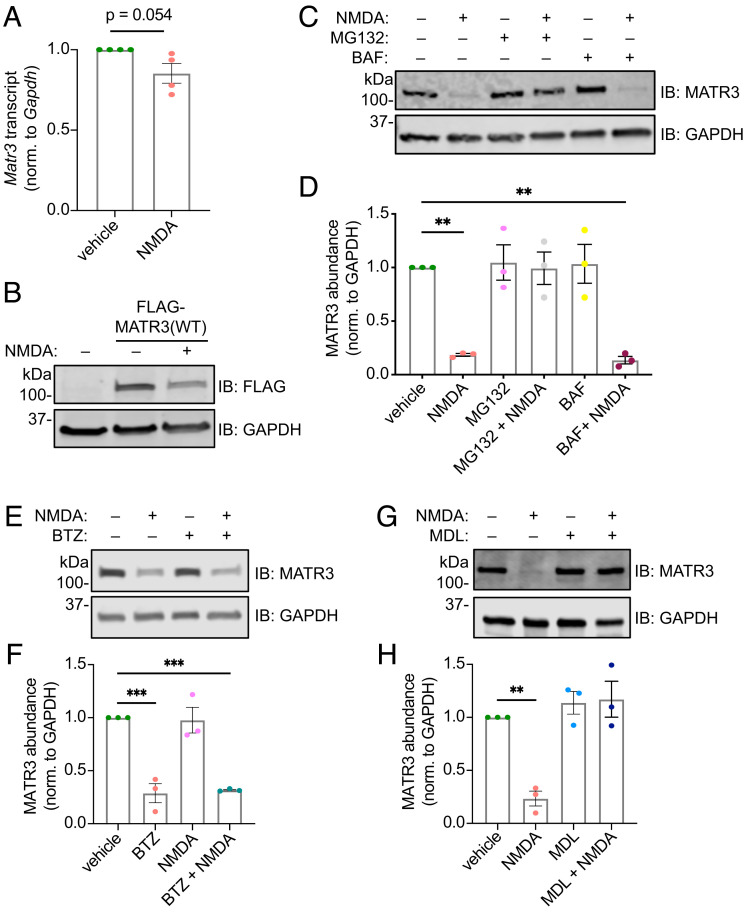
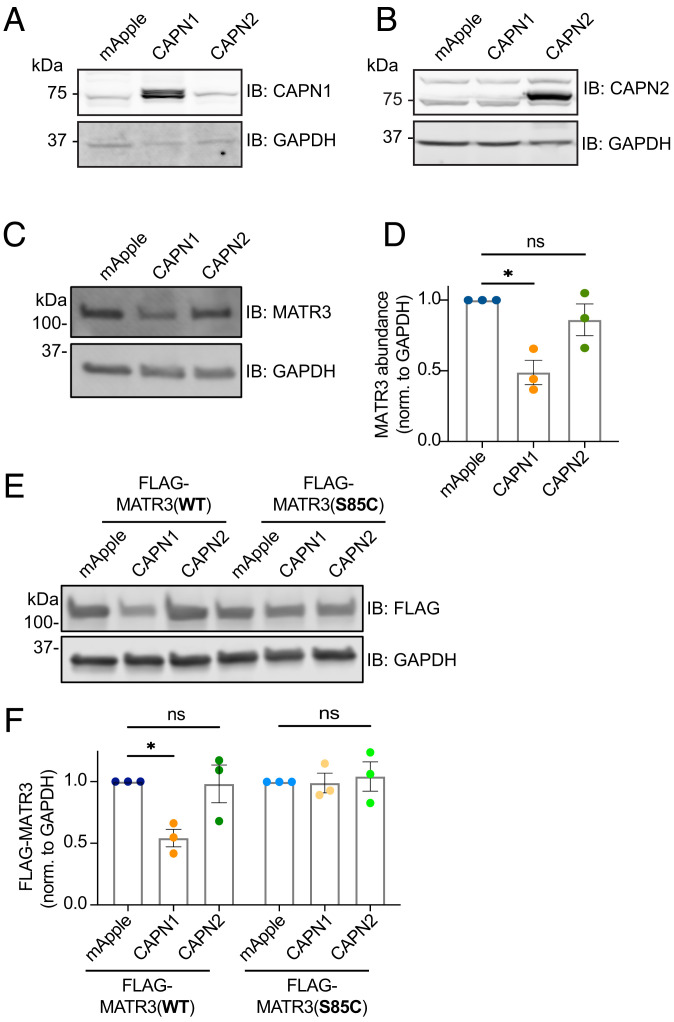
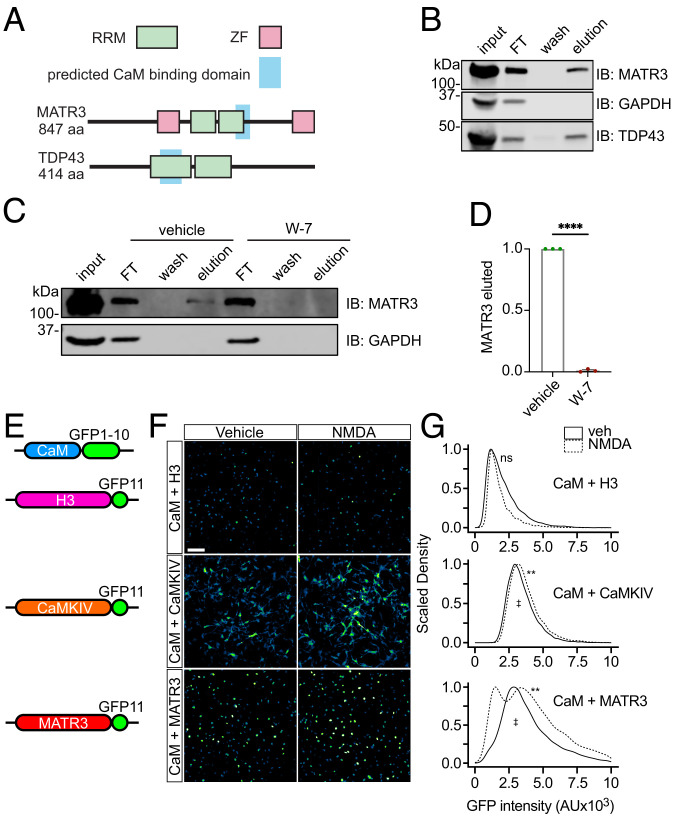
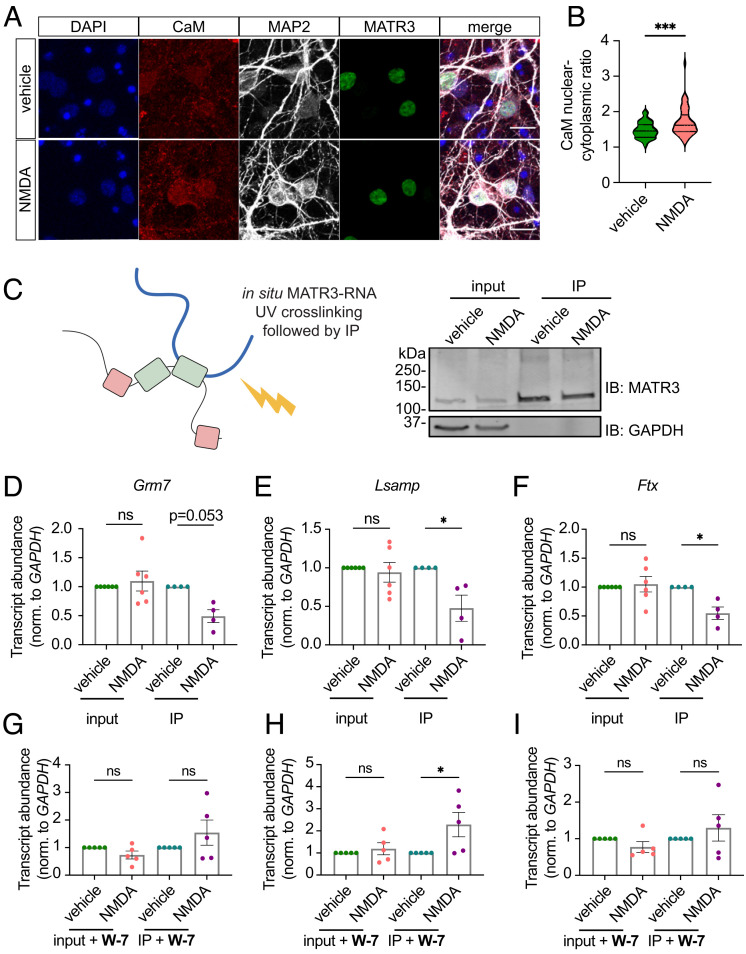
RNA-binding protein (RBP) dysfunction is a fundamental hallmark of amyotrophic lateral sclerosis (ALS) and related neuromuscular disorders. Abnormal neuronal excitability is also a conserved feature in ALS patients and disease models, yet little is known about how activity-dependent processes regulate RBP levels and functions. Mutations in the gene encoding the RBP Matrin 3 (MATR3) cause familial disease, and MATR3 pathology has also been observed in sporadic ALS, suggesting a key role for MATR3 in disease pathogenesis. Here, we show that glutamatergic activity drives MATR3 degradation through an NMDA receptor-, Ca2+-, and calpain-dependent mechanism. The most common pathogenic MATR3 mutation renders it resistant to calpain degradation, suggesting a link between activity-dependent MATR3 regulation and disease. We also demonstrate that Ca2+ regulates MATR3 through a nondegradative process involving the binding of Ca2+/calmodulin to MATR3 and inhibition of its RNA-binding ability. These findings indicate that neuronal activity impacts both the abundance and function of MATR3, underscoring the effect of activity on RBPs and providing a foundation for further study of Ca2+-coupled regulation of RBPs implicated in ALS and related neurological diseases.
Keywords: ALS; FTD; NMDA; RNA binding protein.
Conflict of interest statement
The authors declare no competing interest.
Figures



Similar articles
-
The role of Matrin-3 in physiology and its dysregulation in disease.Biochem Soc Trans. 2024 Jun 26;52(3):961-972. doi: 10.1042/BST20220585. Biochem Soc Trans. 2024. PMID: 38813817 Free PMC article. Review.
-
Matrin 3-dependent neurotoxicity is modified by nucleic acid binding and nucleocytoplasmic localization.Elife. 2018 Jul 17;7:e35977. doi: 10.7554/eLife.35977. Elife. 2018. PMID: 30015619 Free PMC article.
-
Matrin 3 Is a Component of Neuronal Cytoplasmic Inclusions of Motor Neurons in Sporadic Amyotrophic Lateral Sclerosis.Am J Pathol. 2018 Feb;188(2):507-514. doi: 10.1016/j.ajpath.2017.10.007. Epub 2017 Nov 9. Am J Pathol. 2018. PMID: 29128563
-
RNA-recognition motif in Matrin-3 mediates neurodegeneration through interaction with hnRNPM.Acta Neuropathol Commun. 2020 Aug 18;8(1):138. doi: 10.1186/s40478-020-01021-5. Acta Neuropathol Commun. 2020. PMID: 32811564 Free PMC article.
-
Matrin 3 in neuromuscular disease: physiology and pathophysiology.JCI Insight. 2021 Jan 11;6(1):e143948. doi: 10.1172/jci.insight.143948. JCI Insight. 2021. PMID: 33427209 Free PMC article. Review.
Cited by
-
The role of Matrin-3 in physiology and its dysregulation in disease.Biochem Soc Trans. 2024 Jun 26;52(3):961-972. doi: 10.1042/BST20220585. Biochem Soc Trans. 2024. PMID: 38813817 Free PMC article. Review.
-
TRPC4 deletion elicits behavioral defects in sociability by dysregulating expression of microRNA-138-2.iScience. 2023 Dec 2;27(1):108617. doi: 10.1016/j.isci.2023.108617. eCollection 2024 Jan 19. iScience. 2023. PMID: 38188509 Free PMC article.
References
-
- Arai T., et al. , TDP-43 is a component of ubiquitin-positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Biochem. Biophys. Res. Commun. 351, 602–611 (2006). - PubMed
-
- Neumann M., et al. , Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science 314, 130–133 (2006). - PubMed
-
- Kabashi E., et al. , TARDBP mutations in individuals with sporadic and familial amyotrophic lateral sclerosis. Nat. Genet. 40, 572–574 (2008). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
