

Regulated control of virus replication by 4-hydroxytamoxifen-induced splicing
- PMID: 36992923
- PMCID: PMC10040539
- DOI: 10.3389/fmicb.2023.1112580
Regulated control of virus replication by 4-hydroxytamoxifen-induced splicing
Abstract
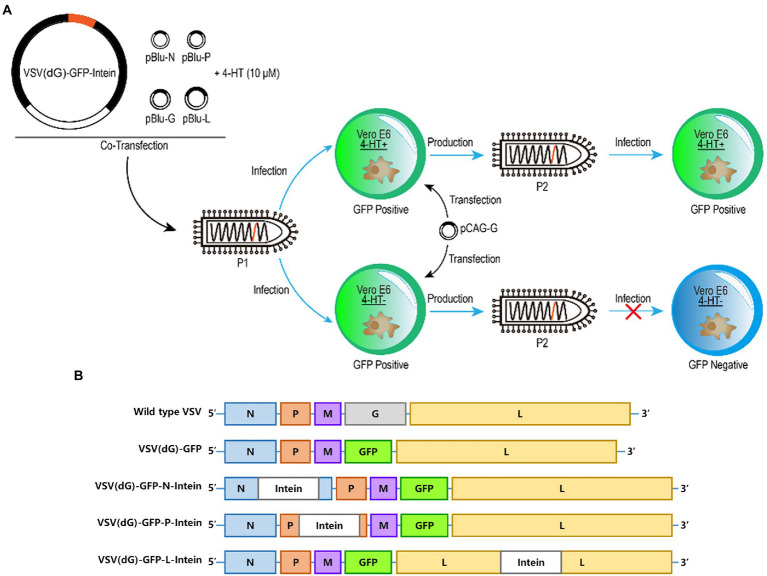
Designing a modified virus that can be controlled to replicate will facilitate the study of pathogenic mechanisms of virus and virus-host interactions. Here, we report a universal switch element that enables precise control of virus replication after exposure to a small molecule. Inteins mediate a traceless protein splicing-ligation process, and we generate a series of modified vesicular stomatitis virus (VSV) with intein insertion into the nucleocapsid, phosphoprotein, or large RNA-dependent RNA polymerase of VSV. Two recombinant VSV, LC599 and LY1744, were screened for intein insertion in the large RNA-dependent RNA polymerase of VSV, and their replication was regulated in a dose-dependent manner with the small molecule 4-hydroxytamoxifen, which induces intein splicing to restore the VSV replication. Furthermore, in the presence of 4-hydroxytamoxifen, the intein-modified VSV LC599 replicated efficiently in an animal model like a prototype of VSV. Thus, we present a simple and highly adaptable tool for regulating virus replication.
Keywords: intein; post-translation regulation; small molecule switch; vesicular stomatitis virus; virus regulation.
Copyright © 2023 Zhao, Wang, Wu, Zhang, Chen, Zhang, Wang, Zhu, Li, Xu, Hou and Chen.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Oligomerization of the Vesicular Stomatitis Virus Phosphoprotein Is Dispensable for mRNA Synthesis but Facilitates RNA Replication.J Virol. 2020 Jun 16;94(13):e00115-20. doi: 10.1128/JVI.00115-20. Print 2020 Jun 16. J Virol. 2020. PMID: 32321813 Free PMC article.
-
Replication and amplification of defective interfering particle RNAs of vesicular stomatitis virus in cells expressing viral proteins from vectors containing cloned cDNAs.J Virol. 1990 Jun;64(6):2948-57. doi: 10.1128/JVI.64.6.2948-2957.1990. J Virol. 1990. PMID: 2159555 Free PMC article.
-
Regulation of protein activity with small-molecule-controlled inteins.Protein Sci. 2005 Feb;14(2):523-32. doi: 10.1110/ps.04996905. Epub 2005 Jan 4. Protein Sci. 2005. PMID: 15632292 Free PMC article.
-
Vesicular stomatitis virus oncolysis of T lymphocytes requires cell cycle entry and translation initiation.J Virol. 2008 Jun;82(12):5735-49. doi: 10.1128/JVI.02601-07. Epub 2008 Apr 16. J Virol. 2008. PMID: 18417567 Free PMC article.
-
Immunovirotherapy Based on Recombinant Vesicular Stomatitis Virus: Where Are We?Front Immunol. 2022 Jun 28;13:898631. doi: 10.3389/fimmu.2022.898631. eCollection 2022. Front Immunol. 2022. PMID: 35837384 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources