

Regulation of adult stem cell quiescence and its functions in the maintenance of tissue integrity
- PMID: 36922629
- PMCID: PMC10725182
- DOI: 10.1038/s41580-022-00568-6
Regulation of adult stem cell quiescence and its functions in the maintenance of tissue integrity
Abstract
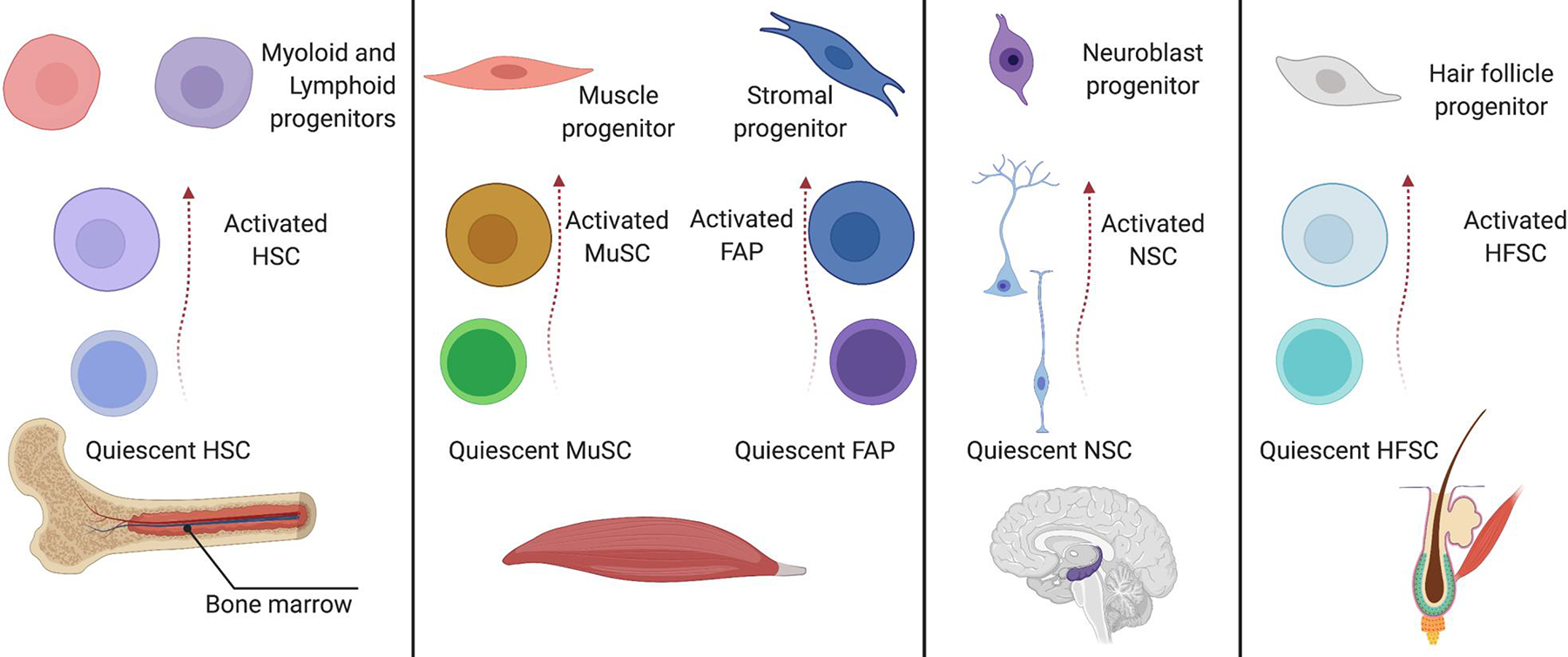
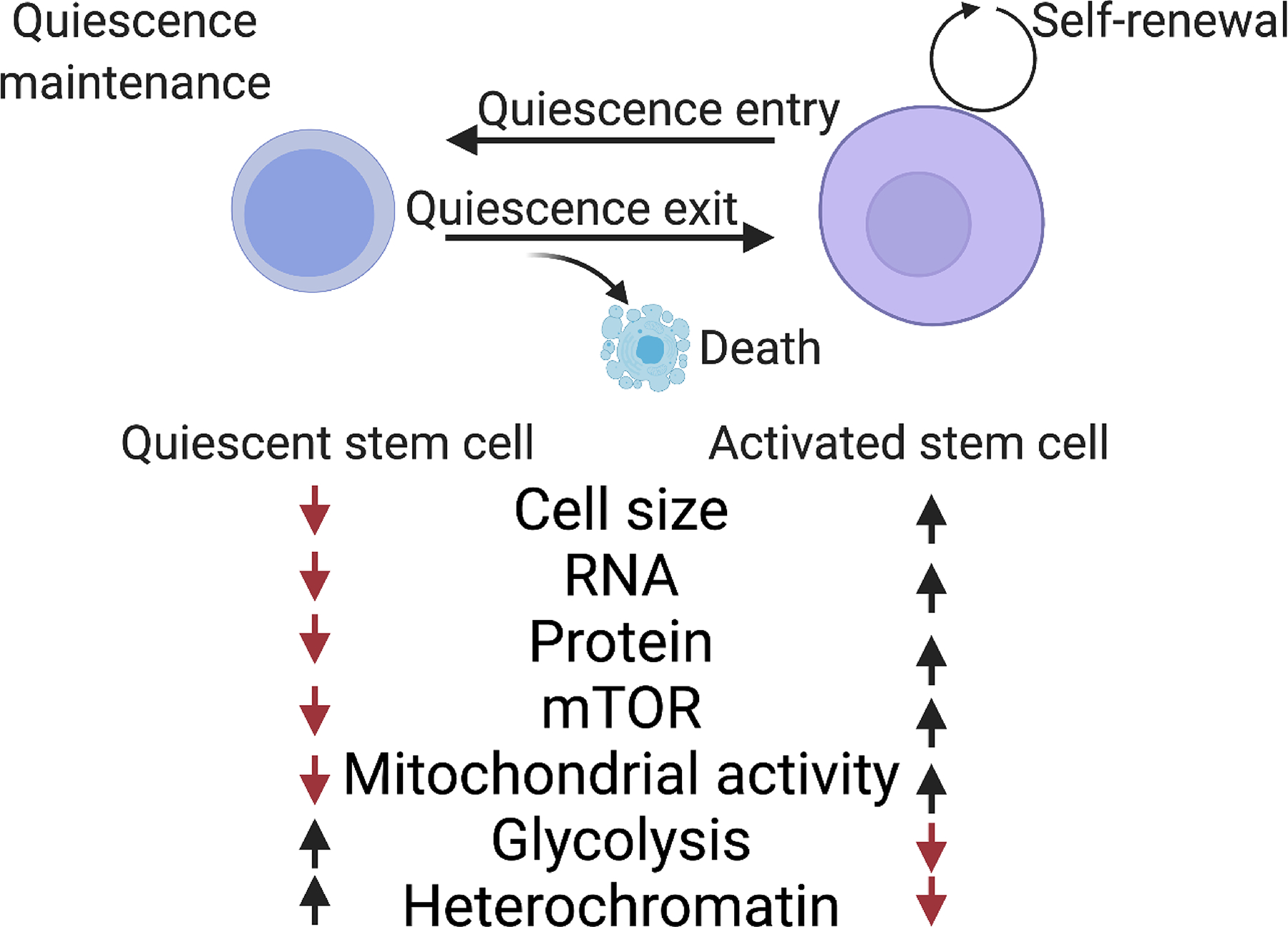
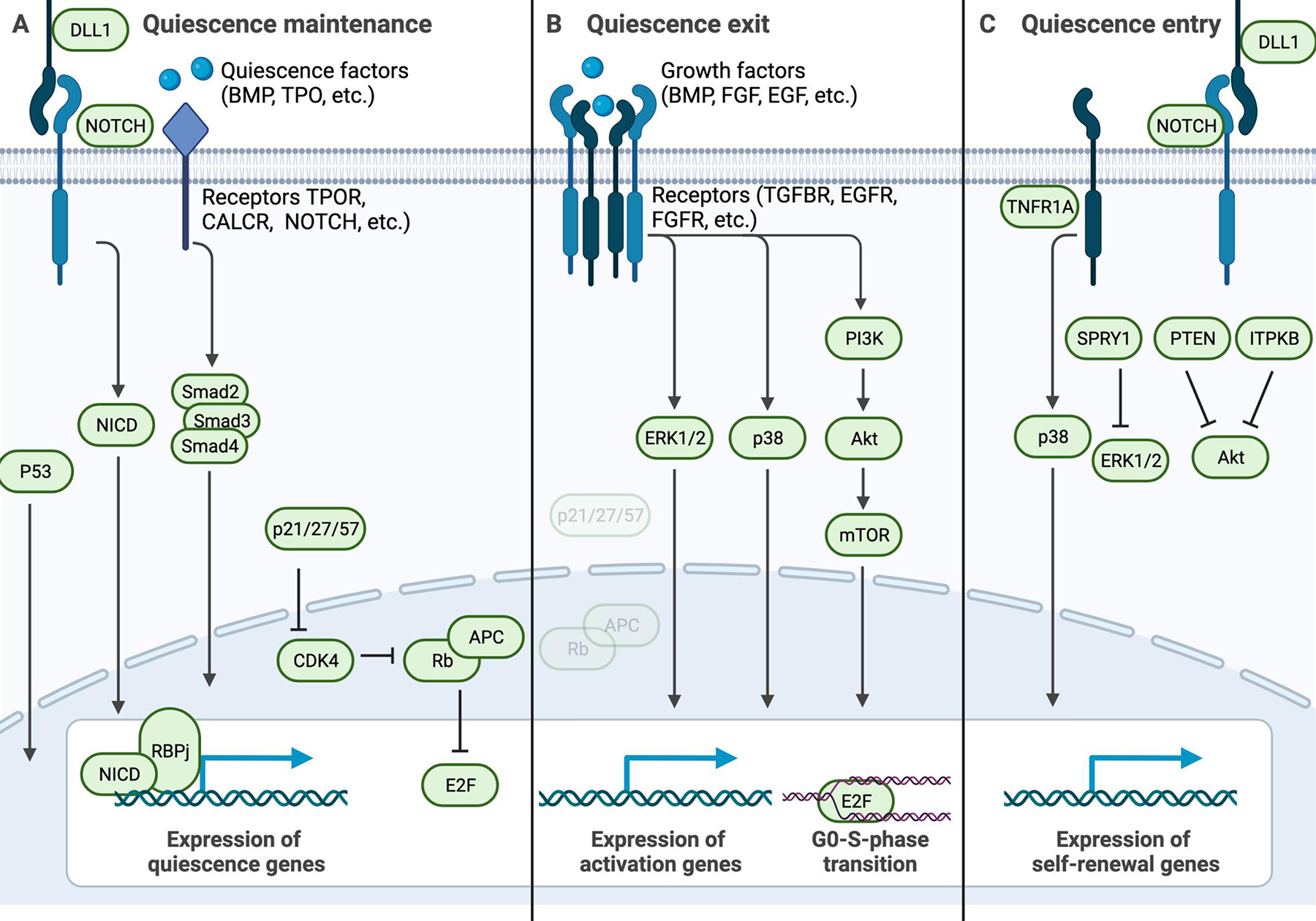
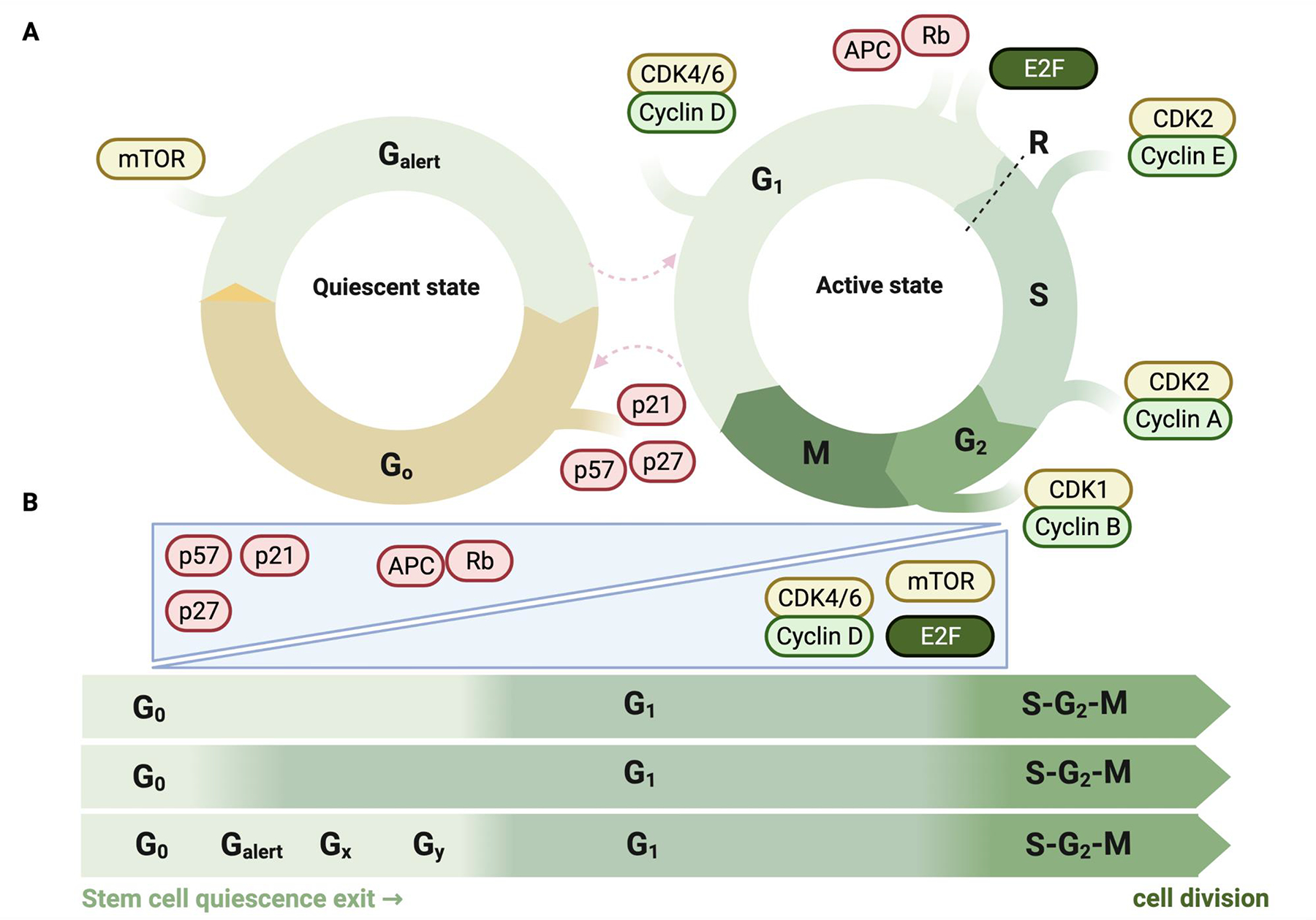
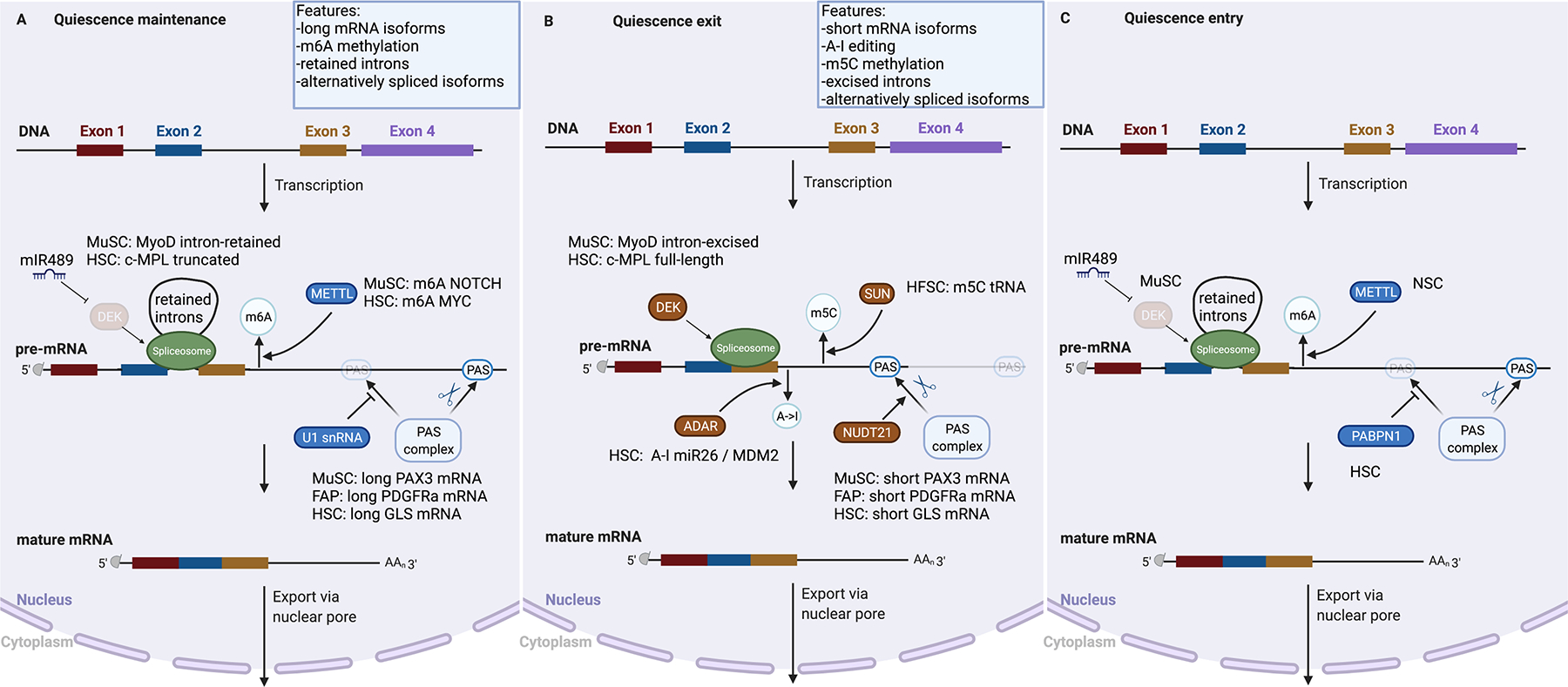
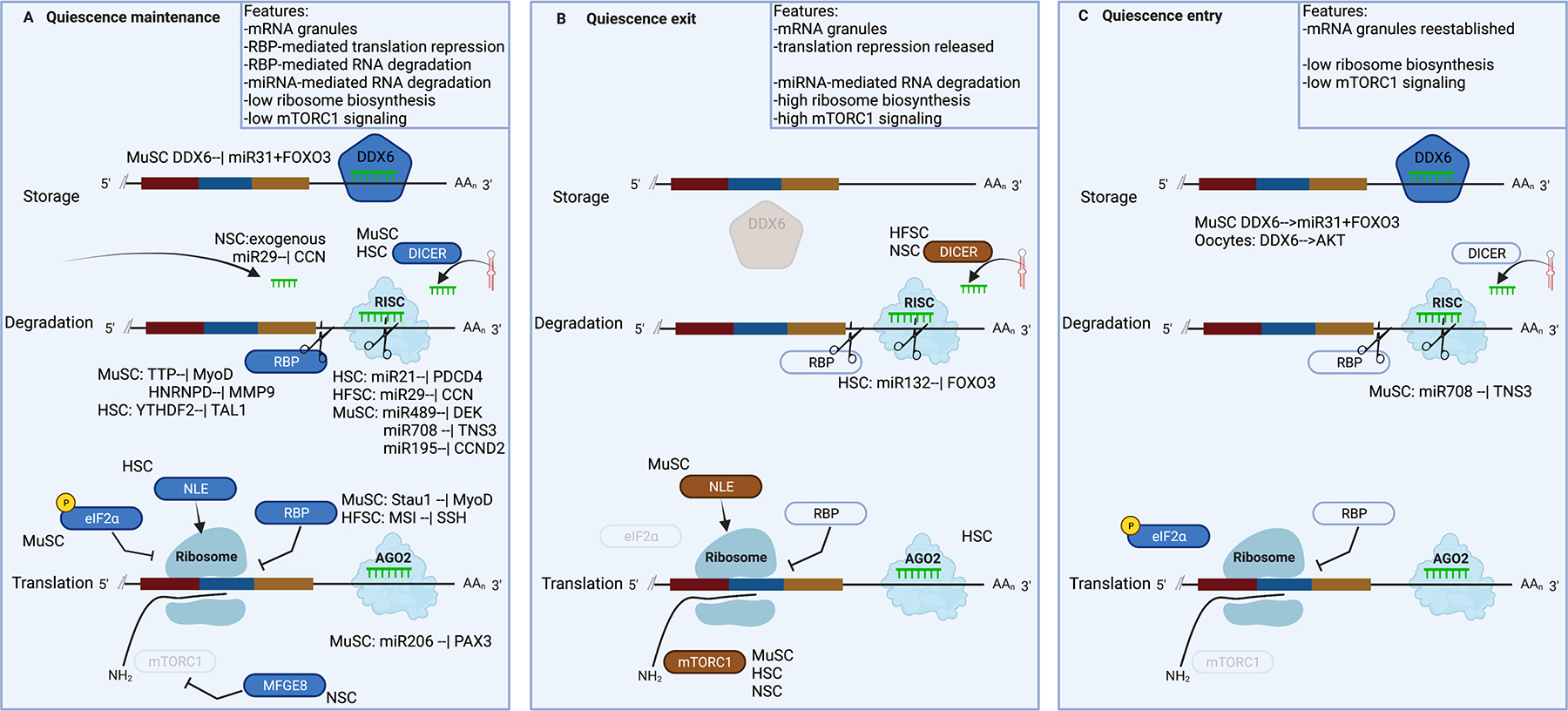
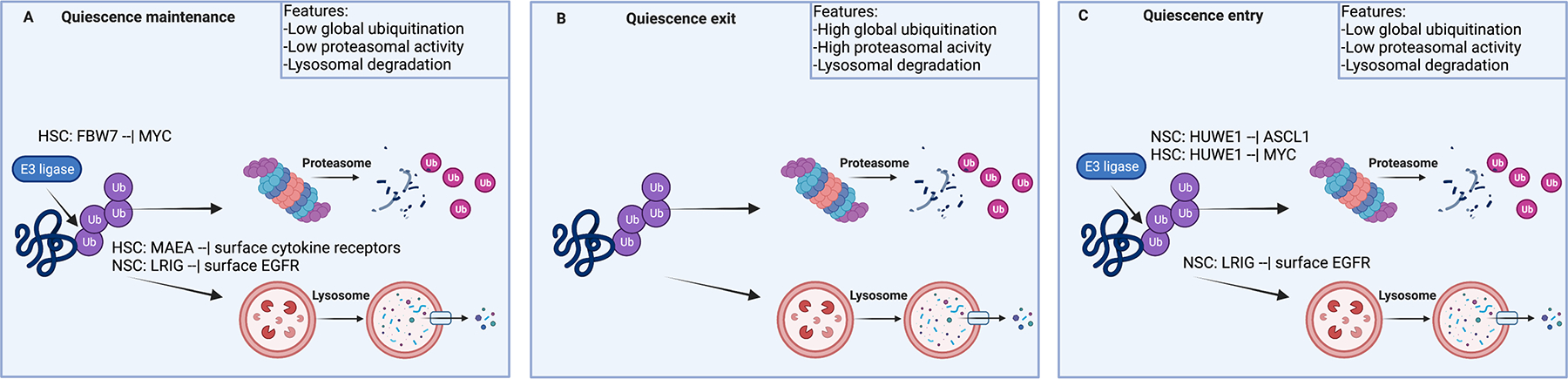
Adult stem cells are important for mammalian tissues, where they act as a cell reserve that supports normal tissue turnover and can mount a regenerative response following acute injuries. Quiescent stem cells are well established in certain tissues, such as skeletal muscle, brain, and bone marrow. The quiescent state is actively controlled and is essential for long-term maintenance of stem cell pools. In this Review, we discuss the importance of maintaining a functional pool of quiescent adult stem cells, including haematopoietic stem cells, skeletal muscle stem cells, neural stem cells, hair follicle stem cells, and mesenchymal stem cells such as fibro-adipogenic progenitors, to ensure tissue maintenance and repair. We discuss the molecular mechanisms that regulate the entry into, maintenance of, and exit from the quiescent state in mice. Recent studies revealed that quiescent stem cells have a discordance between RNA and protein levels, indicating the importance of post-transcriptional mechanisms, such as alternative polyadenylation, alternative splicing, and translation repression, in the control of stem cell quiescence. Understanding how these mechanisms guide stem cell function during homeostasis and regeneration has important implications for regenerative medicine.
© 2023. Springer Nature Limited.
Conflict of interest statement
Competing interests statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Dormancy and quiescence of skeletal muscle stem cells.Results Probl Cell Differ. 2015;56:215-35. doi: 10.1007/978-3-662-44608-9_10. Results Probl Cell Differ. 2015. PMID: 25344673 Review.
-
Quiescence Entry, Maintenance, and Exit in Adult Stem Cells.Int J Mol Sci. 2019 May 1;20(9):2158. doi: 10.3390/ijms20092158. Int J Mol Sci. 2019. PMID: 31052375 Free PMC article. Review.
-
The mini-IDLE 3D biomimetic culture assay enables interrogation of mechanisms governing muscle stem cell quiescence and niche repopulation.Elife. 2022 Dec 20;11:e81738. doi: 10.7554/eLife.81738. Elife. 2022. PMID: 36537758 Free PMC article.
-
Concise Review: Mechanisms of Quiescent Hair Follicle Stem Cell Regulation.Stem Cells. 2017 Dec;35(12):2323-2330. doi: 10.1002/stem.2696. Epub 2017 Sep 23. Stem Cells. 2017. PMID: 28856849 Free PMC article. Review.
-
Concise Review: Quiescence in Adult Stem Cells: Biological Significance and Relevance to Tissue Regeneration.Stem Cells. 2015 Oct;33(10):2903-12. doi: 10.1002/stem.2056. Epub 2015 Jul 27. Stem Cells. 2015. PMID: 26075660 Review.
Cited by
-
Nano-Topographically Guided, Biomineralized, 3D-Printed Polycaprolactone Scaffolds with Urine-Derived Stem Cells for Promoting Bone Regeneration.Pharmaceutics. 2024 Jan 31;16(2):204. doi: 10.3390/pharmaceutics16020204. Pharmaceutics. 2024. PMID: 38399258 Free PMC article.
-
Senescent Cells: Dual Implications on the Retinal Vascular System.Cells. 2023 Sep 23;12(19):2341. doi: 10.3390/cells12192341. Cells. 2023. PMID: 37830555 Free PMC article. Review.
-
Integrative multi-omics analysis unveils stemness-associated molecular subtypes in prostate cancer and pan-cancer: prognostic and therapeutic significance.J Transl Med. 2023 Nov 7;21(1):789. doi: 10.1186/s12967-023-04683-6. J Transl Med. 2023. PMID: 37936202 Free PMC article.
-
Remyelinating Drugs at a Crossroad: How to Improve Clinical Efficacy and Drug Screenings.Cells. 2024 Aug 8;13(16):1326. doi: 10.3390/cells13161326. Cells. 2024. PMID: 39195216 Free PMC article. Review.
-
The Epigenetic Regulation of Quiescent in Stem Cells.Glob Med Genet. 2023 Nov 22;10(4):339-344. doi: 10.1055/s-0043-1777072. eCollection 2023 Dec. Glob Med Genet. 2023. PMID: 38025190 Free PMC article. Review.
References
-
- Orford KW & Scadden DT Deconstructing stem cell self-renewal: Genetic insights into cell-cycle regulation. Nat. Rev. Genet. 9, 115–128 (2008). - PubMed
-
- Ramalho-Santos M & Willenbring H On the Origin of the Term ‘Stem Cell’. Cell Stem Cell 1, 35–38 (2007). - PubMed
-
- Barker N, Bartfeld S & Clevers H Tissue-resident adult stem cell populations of rapidly self-renewing organs. Cell Stem Cell 7, 656–670 (2010). - PubMed
