

Regulation of carotenoid and flavonoid biosynthetic pathways in Lactuca sativa var capitate L. in protected cultivation
- PMID: 36866364
- PMCID: PMC9971571
- DOI: 10.3389/fpls.2023.1124750
Regulation of carotenoid and flavonoid biosynthetic pathways in Lactuca sativa var capitate L. in protected cultivation
Abstract
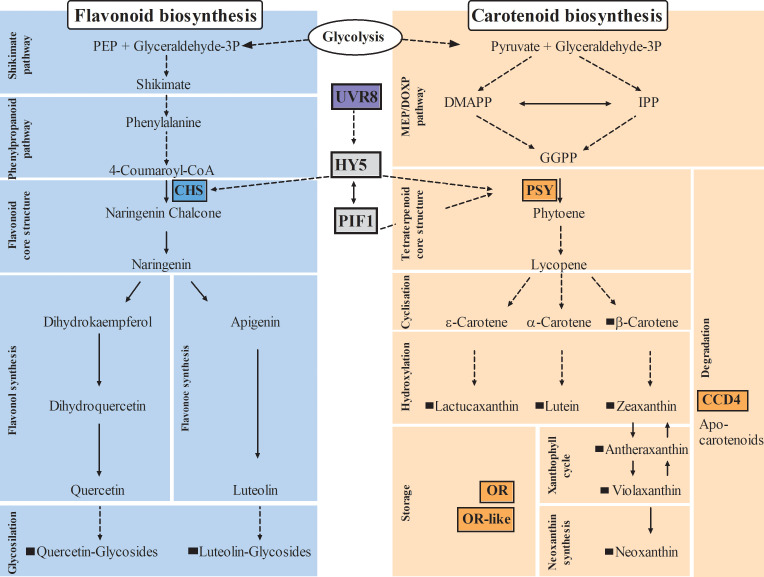
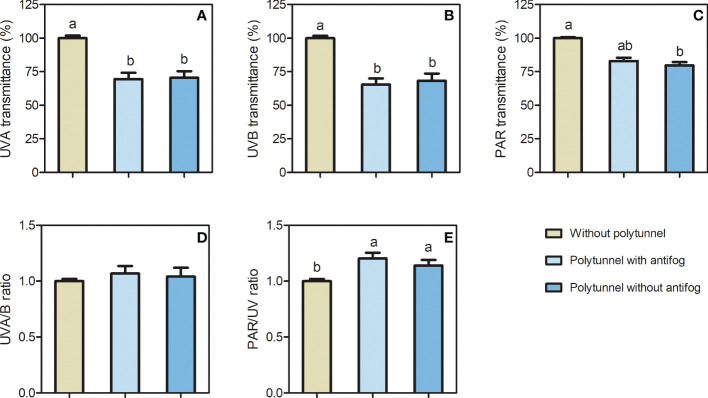
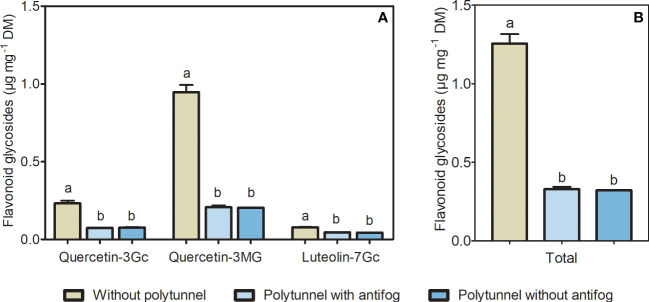
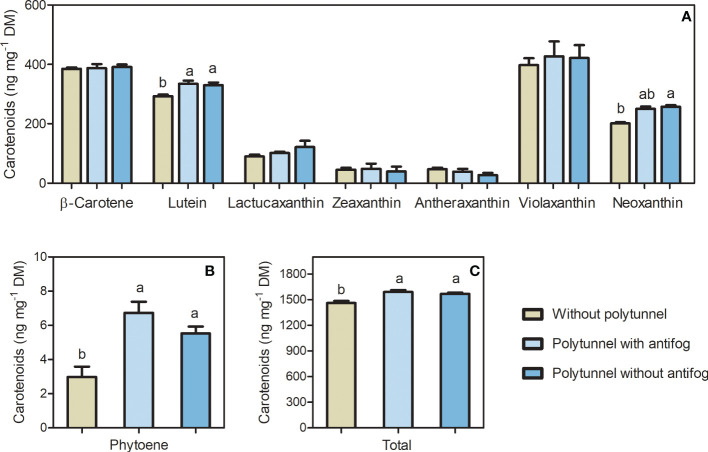
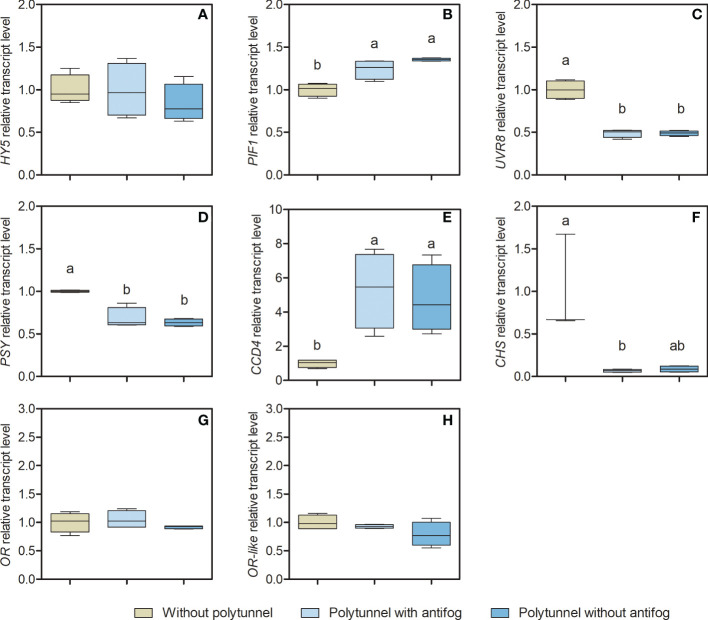
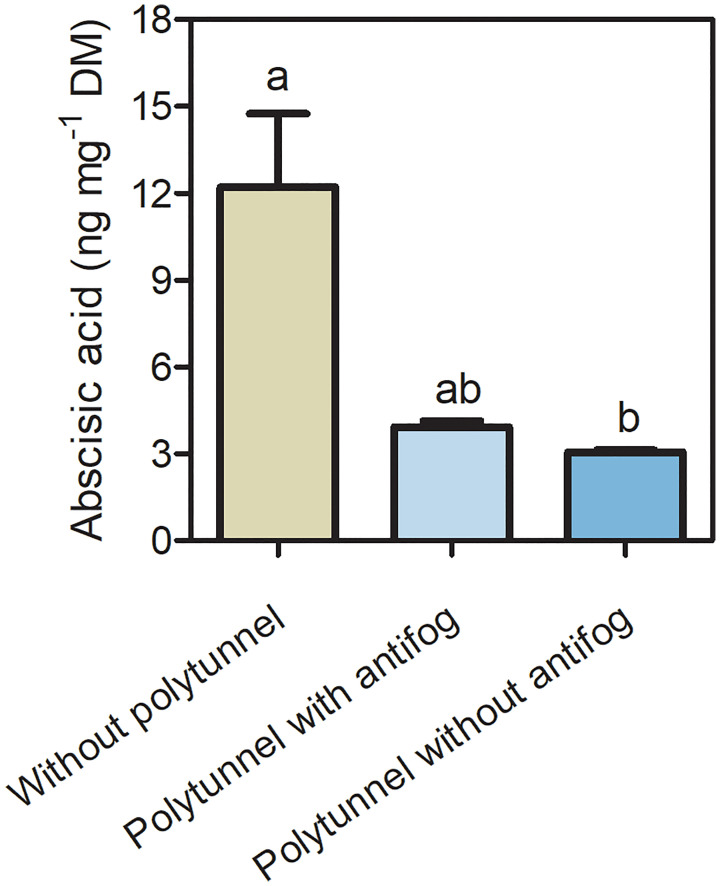
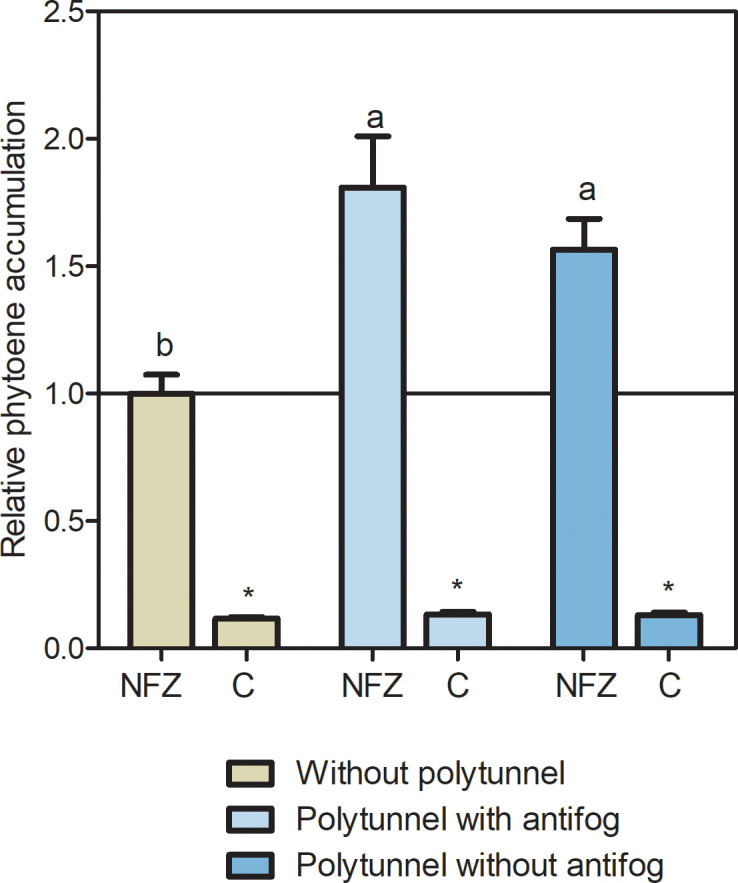
In the face of a growing world population and limited land, there is an urgent demand for higher productivity of food crops, and cultivation systems must be adapted to future needs. Sustainable crop production should aim for not only high yields, but also high nutritional values. In particular, the consumption of bioactive compounds such as carotenoids and flavonoids is associated with a reduced incidence of non-transmissible diseases. Modulating environmental conditions by improving cultivation systems can lead to the adaption of plant metabolisms and the accumulation of bioactive compounds. The present study investigates the regulation of carotenoid and flavonoid metabolisms in lettuce (Lactuca sativa var capitate L.) grown in a protected environment (polytunnels) compared to plants grown without polytunnels. Carotenoid, flavonoid and phytohormone (ABA) contents were determined using HPLC-MS and transcript levels of key metabolic genes were analyzed by RT-qPCR. In this study, we observed inverse contents of flavonoids and carotenoids in lettuce grown without or under polytunnels. Flavonoid contents on a total and individual level were significantly lower, while total carotenoid content was higher in lettuce plants grown under polytunnels compared to without. However, the adaptation was specific to the level of individual carotenoids. For instance, the accumulation of the main carotenoids lutein and neoxanthin was induced while the β-carotene content remained unchanged. In addition, our findings suggest that the flavonoid content of lettuce depends on transcript levels of the key biosynthetic enzyme, which is modulated by UV light. A regulatory influence can be assumed based on the relation between the concentration of the phytohormone ABA and the flavonoid content in lettuce. In contrast, the carotenoid content is not reflected in transcript levels of the key enzyme of either the biosynthetic or the degradation pathway. Nevertheless, the carotenoid metabolic flux determined using norflurazon was higher in lettuce grown under polytunnels, suggesting posttranscriptional regulation of carotenoid accumulation, which should be an integral part of future studies. Therefore, a balance needs to be found between the individual environmental factors, including light and temperature, in order to optimize the carotenoid or flavonoid contents and to obtain nutritionally highly valuable crops in protected cultivation.
Keywords: UV; bioactive compounds; carotenoid; crop cultivation; flavonoid; greenhouse; lettuce; metabolism.
Copyright © 2023 Harbart, Frede, Fitzner and Baldermann.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Regulation of Carotenoid Biosynthesis and Degradation in Lettuce (Lactuca sativa L.) from Seedlings to Harvest.Int J Mol Sci. 2023 Jun 18;24(12):10310. doi: 10.3390/ijms241210310. Int J Mol Sci. 2023. PMID: 37373458 Free PMC article.
-
Changes in the flavonoid and phenolic acid contents and antioxidant activity of red leaf lettuce (Lollo Rosso) due to cultivation under plastic films varying in ultraviolet transparency.J Agric Food Chem. 2007 Dec 12;55(25):10168-72. doi: 10.1021/jf071570m. Epub 2007 Nov 15. J Agric Food Chem. 2007. PMID: 18001028
-
Carotenoid metabolism during bilberry (Vaccinium myrtillus L.) fruit development under different light conditions is regulated by biosynthesis and degradation.BMC Plant Biol. 2016 Apr 21;16:95. doi: 10.1186/s12870-016-0785-5. BMC Plant Biol. 2016. PMID: 27098458 Free PMC article.
-
Regulation of Carotenoid Biosynthesis During Fruit Development.Subcell Biochem. 2016;79:161-98. doi: 10.1007/978-3-319-39126-7_6. Subcell Biochem. 2016. PMID: 27485222 Review.
-
A multifocal approach towards understanding the complexities of carotenoid biosynthesis and accumulation in rice grains.Brief Funct Genomics. 2020 Jul 29;19(4):324-335. doi: 10.1093/bfgp/elaa007. Brief Funct Genomics. 2020. PMID: 32240289 Review.
Cited by
-
Regulation of Carotenoid Biosynthesis and Degradation in Lettuce (Lactuca sativa L.) from Seedlings to Harvest.Int J Mol Sci. 2023 Jun 18;24(12):10310. doi: 10.3390/ijms241210310. Int J Mol Sci. 2023. PMID: 37373458 Free PMC article.
-
The effect of LED light quality on the carotenoid metabolism and related gene expression in the genus Brassica.BMC Plant Biol. 2023 Jun 21;23(1):328. doi: 10.1186/s12870-023-04326-4. BMC Plant Biol. 2023. PMID: 37340342 Free PMC article.
References
-
- Abdallah S.B., Aung B., Amyot L., Lalin I., Lachâal M., Karray-Bouraoui N., et al. . (2016). Salt stress (NaCl) affects plant growth and branch pathways of carotenoid and flavonoid biosyntheses in solanum nigrum. Acta physiologiae plantarum 38, 1–13. doi: 10.1007/s11738-016-2096-8 - DOI
-
- Assumpção C. F., Assis R. Q., Hermes Poletto V. S., Castagna A., Ranieri A., Neugart S., et al. . (2019). Application of supplemental UV-b radiation in pre-harvest to enhance health-promoting compounds accumulation in green and red lettuce. J. Food Process. Preservation 43, e14213. doi: 10.1111/jfpp.14213 - DOI
Grants and funding
LinkOut - more resources
Full Text Sources