

Beyond apoptosis: evidence of other regulated cell death pathways in the ovary throughout development and life
- PMID: 36857094
- PMCID: PMC10320496
- DOI: 10.1093/humupd/dmad005
Beyond apoptosis: evidence of other regulated cell death pathways in the ovary throughout development and life
Abstract
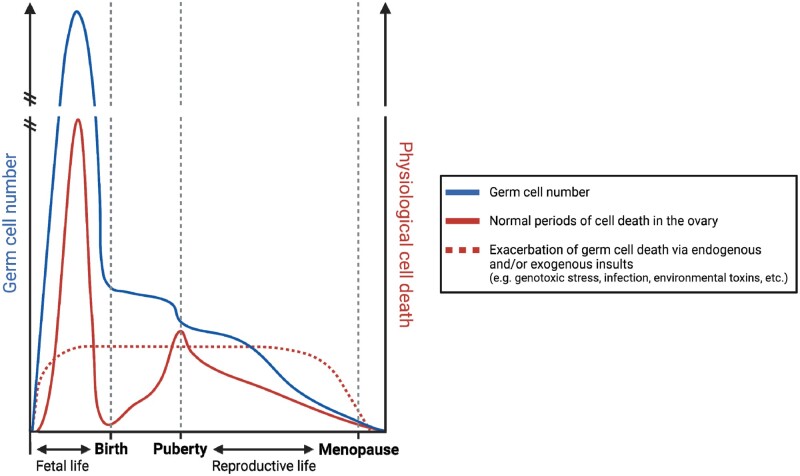
Background: Regulated cell death is a fundamental component of numerous physiological processes; spanning from organogenesis in utero, to normal cell turnover during adulthood, as well as the elimination of infected or damaged cells throughout life. Quality control through regulation of cell death pathways is particularly important in the germline, which is responsible for the generation of offspring. Women are born with their entire supply of germ cells, housed in functional units known as follicles. Follicles contain an oocyte, as well as specialized somatic granulosa cells essential for oocyte survival. Follicle loss-via regulated cell death-occurs throughout follicle development and life, and can be accelerated following exposure to various environmental and lifestyle factors. It is thought that the elimination of damaged follicles is necessary to ensure that only the best quality oocytes are available for reproduction.
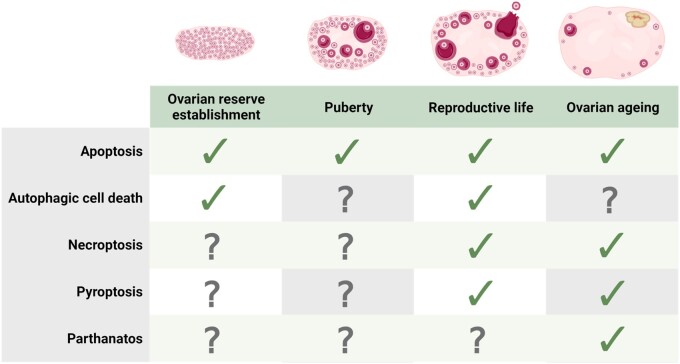
Objective and rationale: Understanding the precise factors involved in triggering and executing follicle death is crucial to uncovering how follicle endowment is initially determined, as well as how follicle number is maintained throughout puberty, reproductive life, and ovarian ageing in women. Apoptosis is established as essential for ovarian homeostasis at all stages of development and life. However, involvement of other cell death pathways in the ovary is less established. This review aims to summarize the most recent literature on cell death regulators in the ovary, with a particular focus on non-apoptotic pathways and their functions throughout the discrete stages of ovarian development and reproductive life.
Search methods: Comprehensive literature searches were carried out using PubMed and Google Scholar for human, animal, and cellular studies published until August 2022 using the following search terms: oogenesis, follicle formation, follicle atresia, oocyte loss, oocyte apoptosis, regulated cell death in the ovary, non-apoptotic cell death in the ovary, premature ovarian insufficiency, primordial follicles, oocyte quality control, granulosa cell death, autophagy in the ovary, autophagy in oocytes, necroptosis in the ovary, necroptosis in oocytes, pyroptosis in the ovary, pyroptosis in oocytes, parthanatos in the ovary, and parthanatos in oocytes.
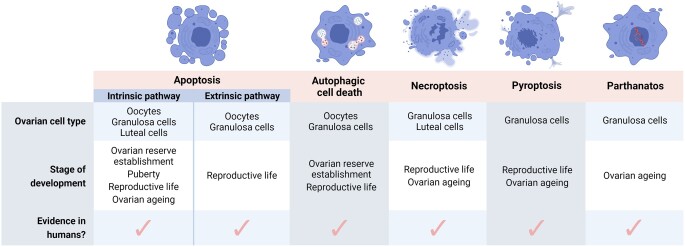
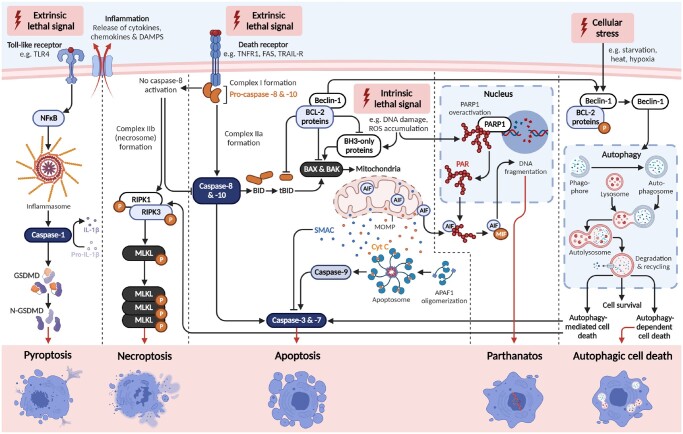
Outcomes: Numerous regulated cell death pathways operate in mammalian cells, including apoptosis, autophagic cell death, necroptosis, and pyroptosis. However, our understanding of the distinct cell death mediators in each ovarian cell type and follicle class across the different stages of life remains the source of ongoing investigation. Here, we highlight recent evidence for the contribution of non-apoptotic pathways to ovarian development and function. In particular, we discuss the involvement of autophagy during follicle formation and the role of autophagic cell death, necroptosis, pyroptosis, and parthanatos during follicle atresia, particularly in response to physiological stressors (e.g. oxidative stress).
Wider implications: Improved knowledge of the roles of each regulated cell death pathway in the ovary is vital for understanding ovarian development, as well as maintenance of ovarian function throughout the lifespan. This information is pertinent not only to our understanding of endocrine health, reproductive health, and fertility in women but also to enable identification of novel fertility preservation targets.
Keywords: apoptosis; autophagy; fertility; granulosa cell; necroptosis; oocyte; ovary; parthanatos; pyroptosis; regulated cell death.
© The Author(s) 2023. Published by Oxford University Press on behalf of European Society of Human Reproduction and Embryology.
Conflict of interest statement
The authors declare no competing financial, or other interests.
Figures
Similar articles
-
Oocyte-somatic cell interactions in the human ovary-novel role of bone morphogenetic proteins and growth differentiation factors.Hum Reprod Update. 2016 Dec;23(1):1-18. doi: 10.1093/humupd/dmw039. Epub 2016 Oct 26. Hum Reprod Update. 2016. PMID: 27797914 Free PMC article. Review.
-
The infant and pubertal human ovary: Balbiani's body-associated VASA expression, immunohistochemical detection of apoptosis-related BCL2 and BAX proteins, and DNA fragmentation.Hum Reprod. 2013 Mar;28(3):698-706. doi: 10.1093/humrep/des453. Epub 2013 Jan 12. Hum Reprod. 2013. PMID: 23315064
-
Role of apoptosis and autophagy in ovarian follicle pool decline in children and women diagnosed with benign or malignant extra-ovarian conditions.Hum Reprod. 2023 Jan 5;38(1):75-88. doi: 10.1093/humrep/deac237. Hum Reprod. 2023. PMID: 36346333
-
Intact fetal ovarian cord formation promotes mouse oocyte survival and development.BMC Dev Biol. 2010 Jan 8;10:2. doi: 10.1186/1471-213X-10-2. BMC Dev Biol. 2010. PMID: 20064216 Free PMC article.
-
[Reconsidering the roles of female germ cells in ovarian development and folliculogenesis].Biol Aujourdhui. 2011;205(4):223-33. doi: 10.1051/jbio/2011022. Epub 2012 Jan 19. Biol Aujourdhui. 2011. PMID: 22251857 Review. French.
Cited by
-
Autophagy and Female Fertility: Mechanisms, Clinical Implications, and Emerging Therapies.Cells. 2024 Aug 14;13(16):1354. doi: 10.3390/cells13161354. Cells. 2024. PMID: 39195244 Free PMC article. Review.
-
Folliculogenesis and steroidogenesis alterations after chronic exposure to a human-relevant mixture of environmental toxicants spare the ovarian reserve in the rabbit model.J Ovarian Res. 2024 Jun 28;17(1):134. doi: 10.1186/s13048-024-01457-6. J Ovarian Res. 2024. PMID: 38943138 Free PMC article.
-
Effect of Serotonin (5-Hydroxytryptamine) on Follicular Development in Porcine.Int J Mol Sci. 2024 Sep 4;25(17):9596. doi: 10.3390/ijms25179596. Int J Mol Sci. 2024. PMID: 39273540 Free PMC article.
-
Premature ovarian insufficiency: a review on the role of oxidative stress and the application of antioxidants.Front Endocrinol (Lausanne). 2023 Aug 1;14:1172481. doi: 10.3389/fendo.2023.1172481. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 37600717 Free PMC article. Review.
-
Inducers and Inhibitors of Pyroptotic Death of Granulosa Cells in Models of Premature Ovarian Insufficiency and Polycystic Ovary Syndrome.Reprod Sci. 2024 Oct;31(10):2972-2992. doi: 10.1007/s43032-024-01643-3. Epub 2024 Jul 18. Reprod Sci. 2024. PMID: 39026050 Free PMC article. Review.
References
-
- Allan CM, Wang Y, Jimenez M, Marshan B, Spaliviero J, Illingworth P, Handelsman DJ.. Follicle-stimulating hormone increases primordial follicle reserve in mature female hypogonadal mice. J Endocrinol 2006;188:549–557. - PubMed
