

The ecl family gene ecl3+ is induced by phosphate starvation and contributes to sexual differentiation in fission yeast
- PMID: 36779416
- PMCID: PMC10038150
- DOI: 10.1242/jcs.260759
The ecl family gene ecl3+ is induced by phosphate starvation and contributes to sexual differentiation in fission yeast
Abstract
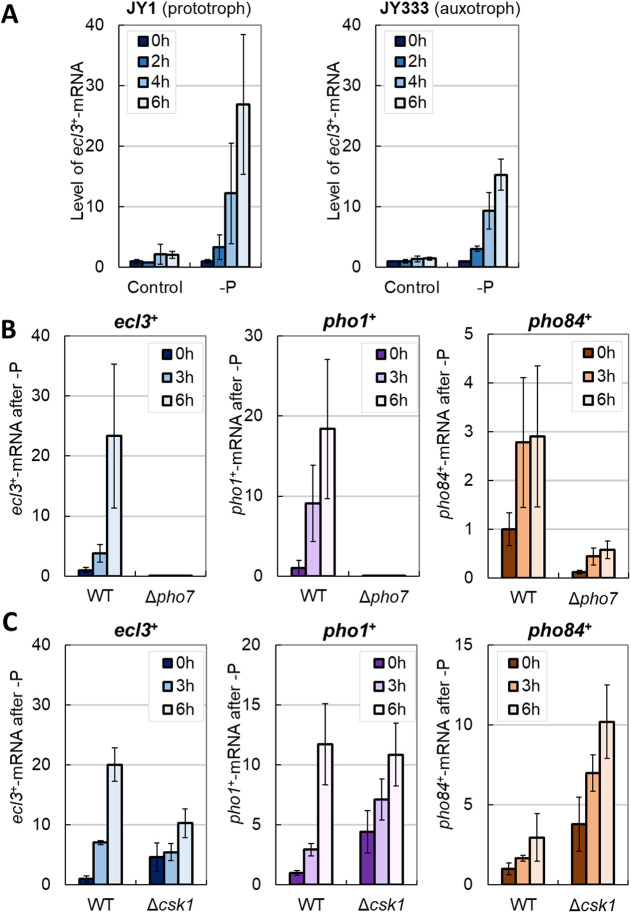
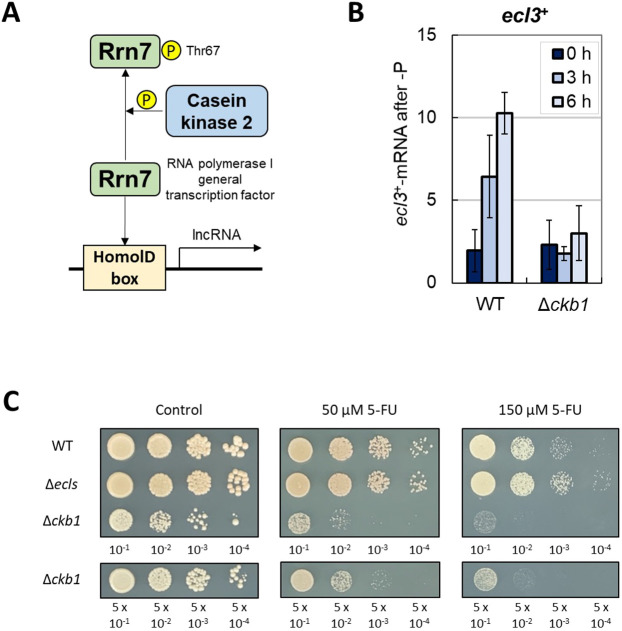
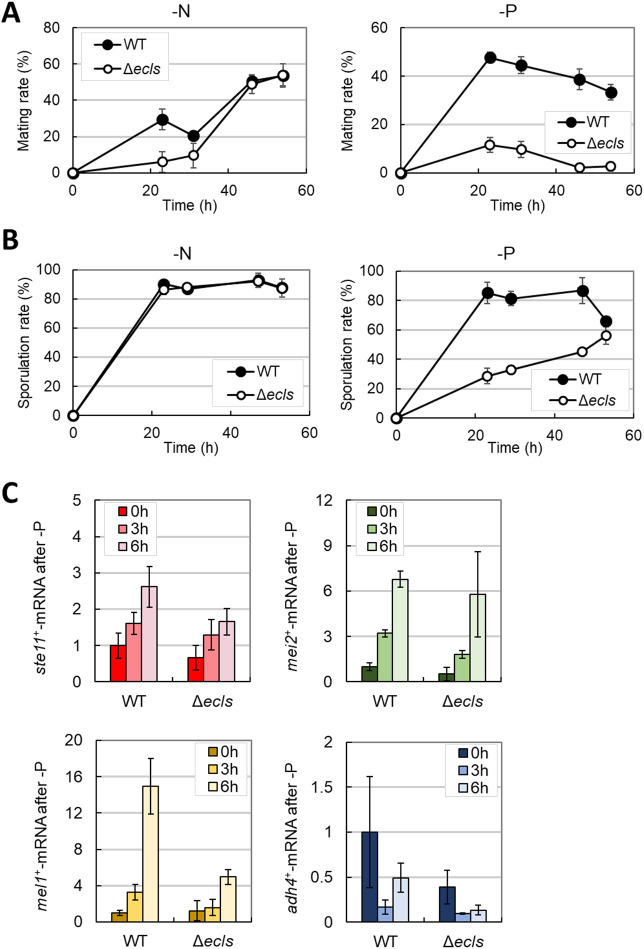
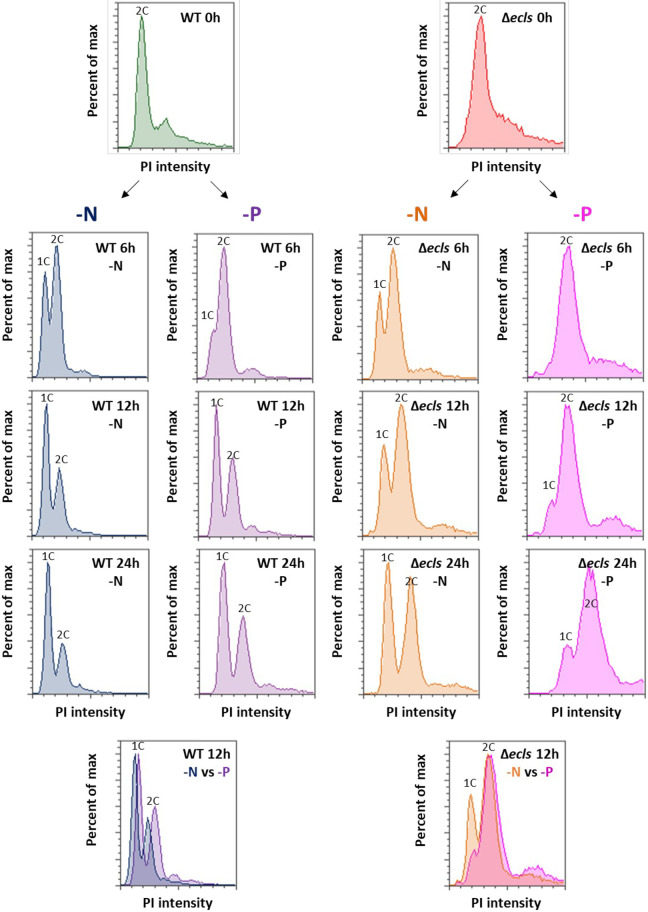
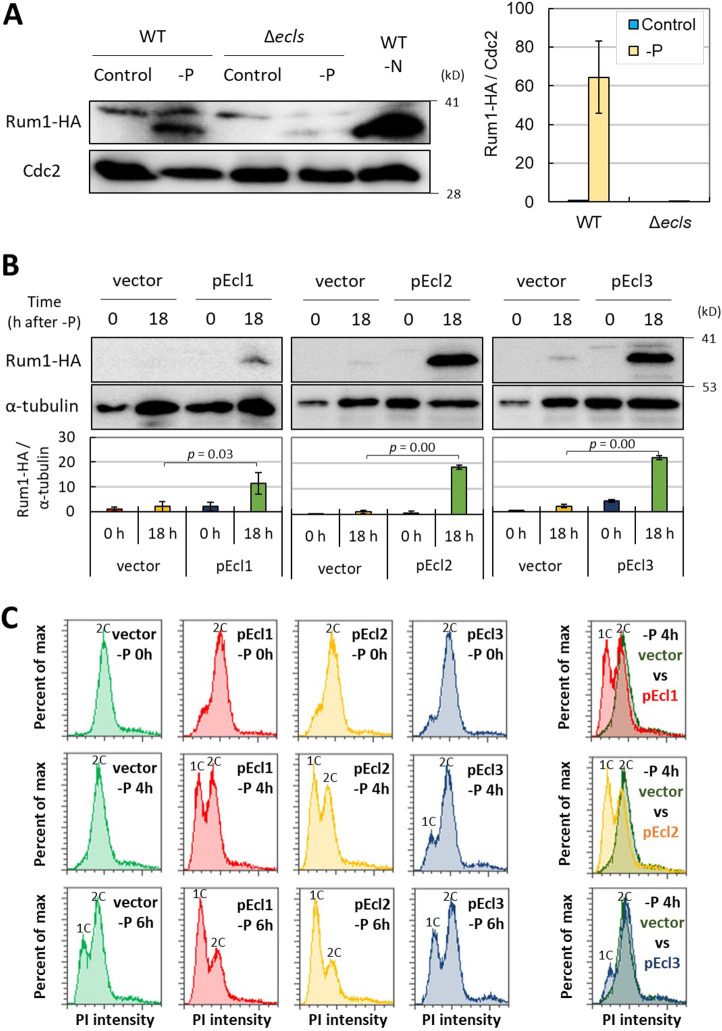
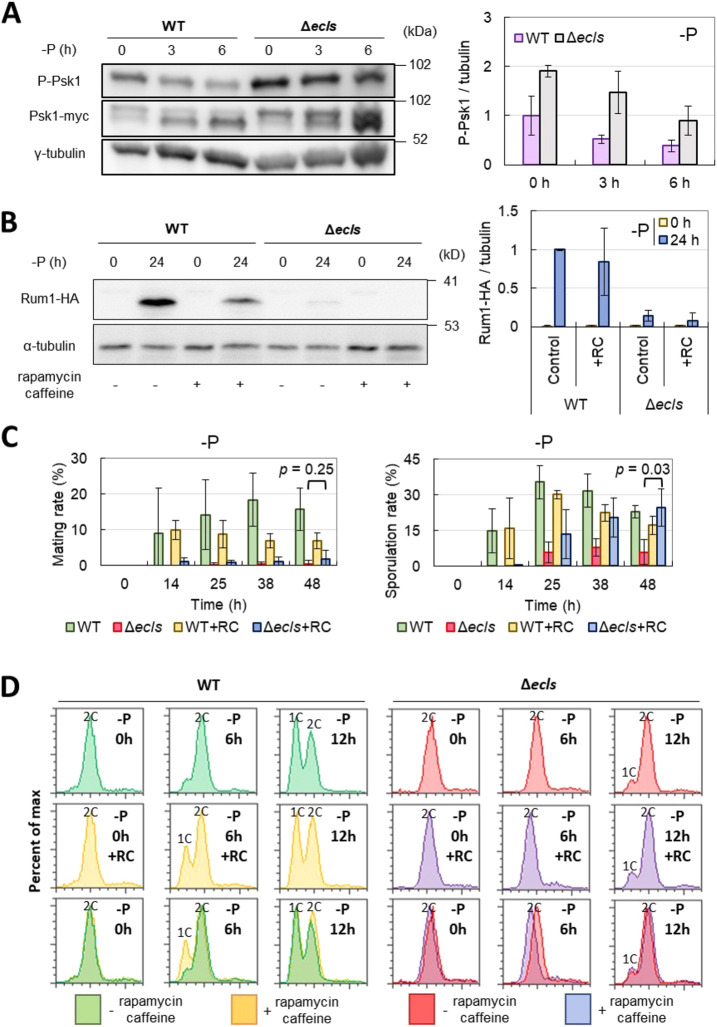
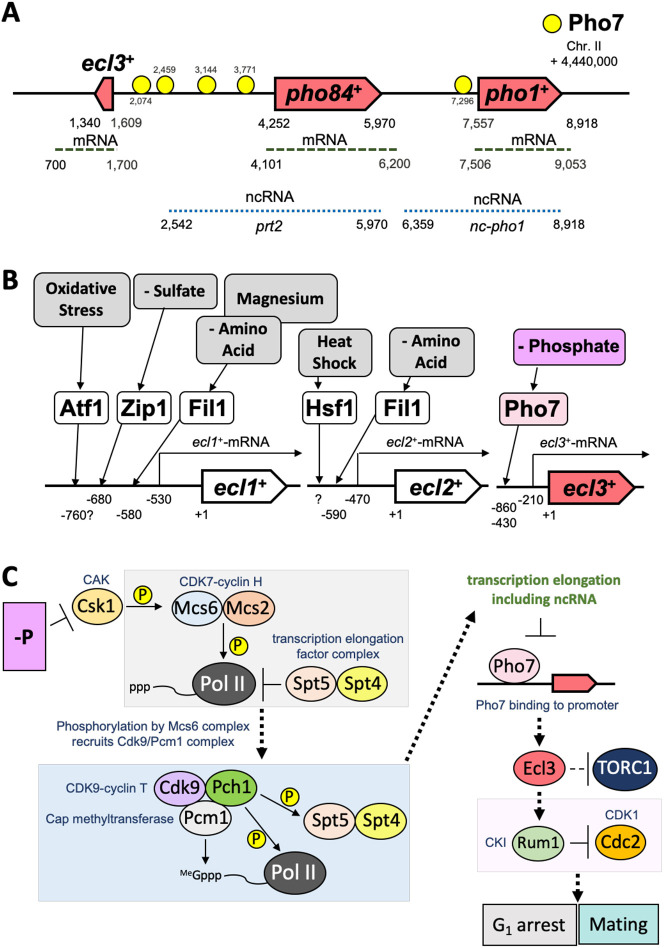
In Schizosaccharomyces pombe, ecl family genes are induced by several signals, such as starvation of various nutrients, including sulfur, amino acids and Mg2+, and environmental stress, including heat or oxidative stress. These genes mediate appropriate cellular responses and contribute to the maintenance of cell viability and induction of sexual differentiation. Although this yeast has three ecl family genes with overlapping functions, any environmental conditions that induce ecl3+ remain unidentified. We demonstrate that ecl3+ is induced by phosphate starvation, similar to its chromosomally neighboring genes, pho1+ and pho84+, which respectively encode an extracellular acid phosphatase and an inorganic phosphate transporter. ecl3+ expression was induced by the transcription factor Pho7 and affected by the cyclin-dependent kinase (CDK)-activating kinase Csk1. Phosphate starvation induced G1 arrest and sexual differentiation via ecl family genes. Biochemical analyses suggested that this G1 arrest was mediated by the stabilization of the CDK inhibitor Rum1, which was dependent on ecl family genes. This study shows that ecl family genes are required for appropriate responses to phosphate starvation and provides novel insights into the diversity and similarity of starvation responses.
Keywords: ecl family genes; Fission yeast; G1 cell cycle arrest; Lifespan; Phosphate signal transduction pathway; Phosphate starvation; Sexual differentiation.
© 2023. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
Similar articles
-
Factors governing the transcriptome changes and chronological lifespan of fission yeast during phosphate starvation.J Biol Chem. 2024 Mar;300(3):105718. doi: 10.1016/j.jbc.2024.105718. Epub 2024 Feb 2. J Biol Chem. 2024. PMID: 38311173 Free PMC article.
-
Genome-wide characterization of the phosphate starvation response in Schizosaccharomyces pombe.BMC Genomics. 2012 Dec 12;13:697. doi: 10.1186/1471-2164-13-697. BMC Genomics. 2012. PMID: 23231582 Free PMC article.
-
Defining the DNA Binding Site Recognized by the Fission Yeast Zn2Cys6 Transcription Factor Pho7 and Its Role in Phosphate Homeostasis.mBio. 2017 Aug 15;8(4):e01218-17. doi: 10.1128/mBio.01218-17. mBio. 2017. PMID: 28811350 Free PMC article.
-
ecl family genes: Factors linking starvation and lifespan extension in Schizosaccharomyces pombe.Mol Microbiol. 2023 Nov;120(5):645-657. doi: 10.1111/mmi.15134. Epub 2023 Jul 31. Mol Microbiol. 2023. PMID: 37525511 Review.
-
Genes affecting the extension of chronological lifespan in Schizosaccharomyces pombe (fission yeast).Mol Microbiol. 2021 Apr;115(4):623-642. doi: 10.1111/mmi.14627. Epub 2020 Nov 3. Mol Microbiol. 2021. PMID: 33064911 Free PMC article. Review.
References
-
- Benjamin, B., Garg, A., Jork, N., Jessen, H. J., Schwer, B. and Shuman, S. (2022). Activities and structure-function analysis of fission yeast inositol pyrophosphate (IPP) kinase-pyrophosphatase Asp1 and its impact on regulation of pho1 gene expression. MBio 13, e0103422. 10.1128/mbio.01034-22 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
