

Animal models of human insomnia
- PMID: 36748845
- PMCID: PMC10404637
- DOI: 10.1111/jsr.13845
Animal models of human insomnia
Abstract
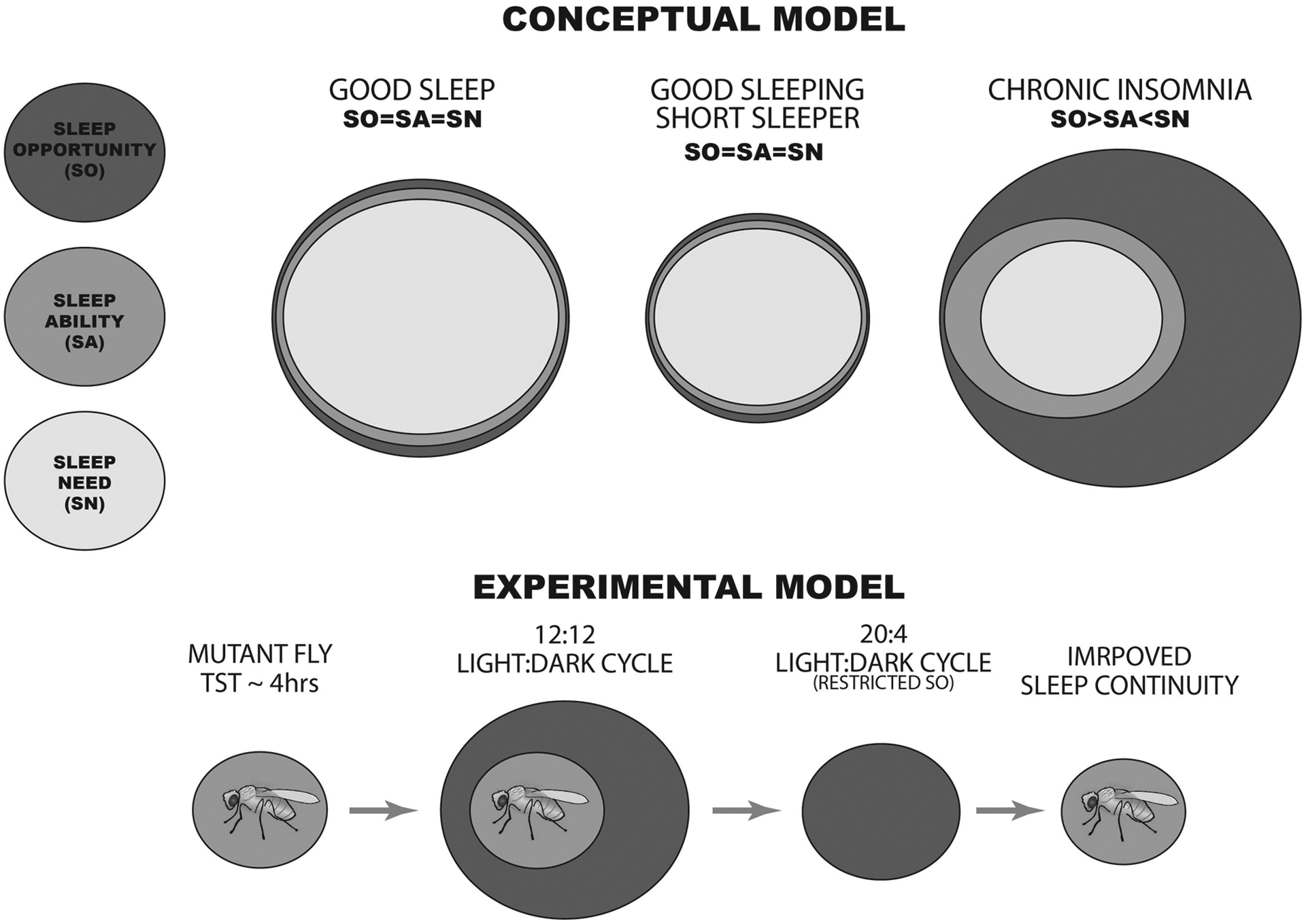
Insomnia disorder (chronic sleep continuity disturbance) is a debilitating condition affecting 5%-10% of the adult population worldwide. To date, researchers have attempted to model insomnia in animals through breeding strategies that create pathologically short-sleeping individuals or with drugs and environmental contexts that directly impose sleeplessness. While these approaches have been invaluable for identifying insomnia susceptibility genes and mapping the neural networks that underpin sleep-wake regulation, they fail to capture concurrently several of the core clinical diagnostic features of insomnia disorder in humans, where sleep continuity disturbance is self-perpetuating, occurs despite adequate sleep opportunity, and is often not accompanied by significant changes in sleep duration or architecture. In the present review, we discuss these issues and then outline ways animal models can be used to develop approaches that are more ecologically valid in their recapitulation of chronic insomnia's natural aetiology and pathophysiology. Conditioning of self-generated sleep loss with these methods promises to create a better understanding of the neuroadaptations that maintain insomnia, including potentially within the infralimbic cortex, a substrate at the crossroads of threat habituation and sleep.
Keywords: fly; infralimbic; insomnia; rodent; sleep disorder; stress perpetuation.
© 2023 European Sleep Research Society.
Figures


Similar articles
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
-
Using Experience Sampling Methodology to Capture Disclosure Opportunities for Autistic Adults.Autism Adulthood. 2023 Dec 1;5(4):389-400. doi: 10.1089/aut.2022.0090. Epub 2023 Dec 12. Autism Adulthood. 2023. PMID: 38116059 Free PMC article.
-
Qualitative evidence synthesis informing our understanding of people's perceptions and experiences of targeted digital communication.Cochrane Database Syst Rev. 2019 Oct 23;10(10):ED000141. doi: 10.1002/14651858.ED000141. Cochrane Database Syst Rev. 2019. PMID: 31643081 Free PMC article.
-
"I've Spent My Whole Life Striving to Be Normal": Internalized Stigma and Perceived Impact of Diagnosis in Autistic Adults.Autism Adulthood. 2023 Dec 1;5(4):423-436. doi: 10.1089/aut.2022.0066. Epub 2023 Dec 12. Autism Adulthood. 2023. PMID: 38116050 Free PMC article.
-
Trends in Surgical and Nonsurgical Aesthetic Procedures: A 14-Year Analysis of the International Society of Aesthetic Plastic Surgery-ISAPS.Aesthetic Plast Surg. 2024 Oct;48(20):4217-4227. doi: 10.1007/s00266-024-04260-2. Epub 2024 Aug 5. Aesthetic Plast Surg. 2024. PMID: 39103642 Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
