

This is a preprint.
DNA supercoiling-induced shapes alter minicircle hydrodynamic properties
- PMID: 36711572
- PMCID: PMC9881935
- DOI: 10.1101/2023.01.04.522747
DNA supercoiling-induced shapes alter minicircle hydrodynamic properties
Update in
-
DNA supercoiling-induced shapes alter minicircle hydrodynamic properties.Nucleic Acids Res. 2023 May 8;51(8):4027-4042. doi: 10.1093/nar/gkad183. Nucleic Acids Res. 2023. PMID: 36971110 Free PMC article.
Abstract
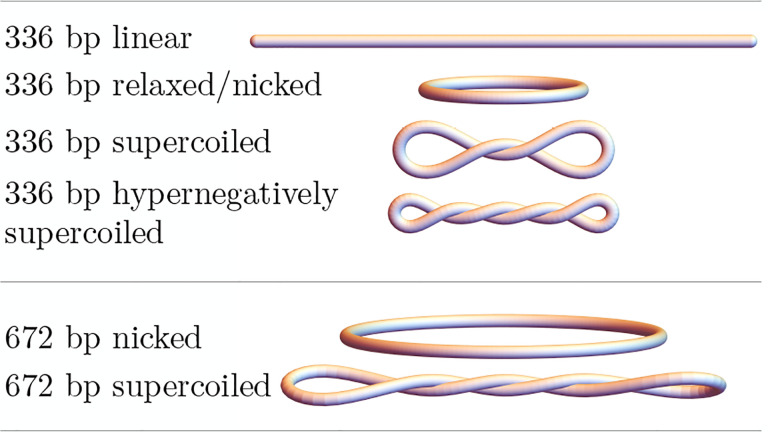
DNA in cells is organized in negatively supercoiled loops. The resulting torsional and bending strain allows DNA to adopt a surprisingly wide variety of 3-D shapes. This interplay between negative supercoiling, looping, and shape influences how DNA is stored, replicated, transcribed, repaired, and likely every other aspect of DNA activity. To understand the consequences of negative supercoiling and curvature on the hydrodynamic properties of DNA, we submitted 336 bp and 672 bp DNA minicircles to analytical ultracentrifugation (AUC). We found that the diffusion coefficient, sedimentation coefficient, and the DNA hydrodynamic radius strongly depended on circularity, loop length, and degree of negative supercoiling. Because AUC cannot ascertain shape beyond degree of non-globularity, we applied linear elasticity theory to predict DNA shapes, and combined these with hydrodynamic calculations to interpret the AUC data, with reasonable agreement between theory and experiment. These complementary approaches, together with earlier electron cryotomography data, provide a framework for understanding and predicting the effects of supercoiling on the shape and hydrodynamic properties of DNA.
Conflict of interest statement
Figures

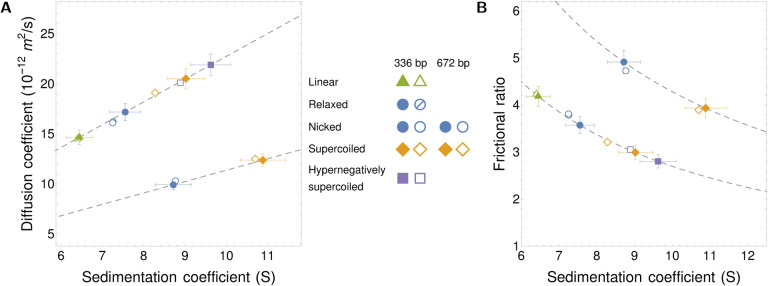
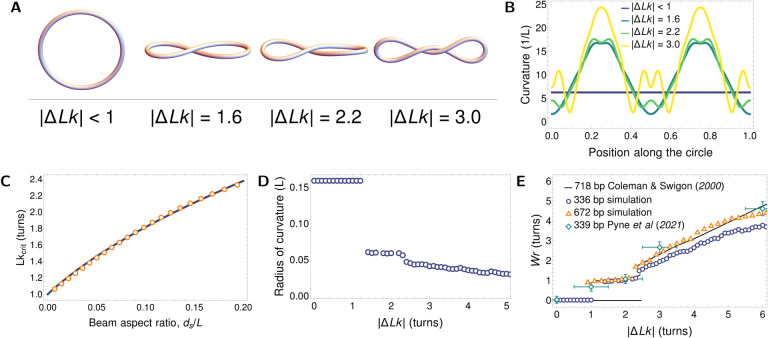
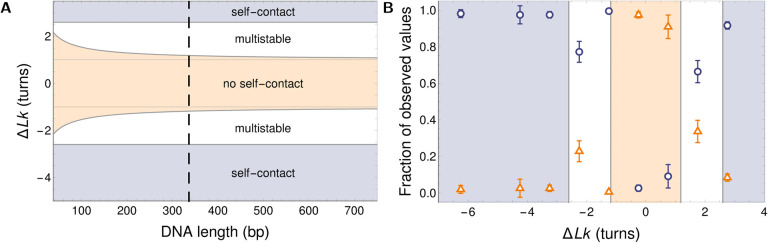
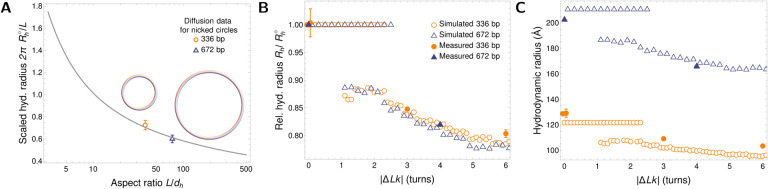
Similar articles
-
DNA supercoiling-induced shapes alter minicircle hydrodynamic properties.Nucleic Acids Res. 2023 May 8;51(8):4027-4042. doi: 10.1093/nar/gkad183. Nucleic Acids Res. 2023. PMID: 36971110 Free PMC article.
-
Supercoiling and looping promote DNA base accessibility and coordination among distant sites.Nat Commun. 2021 Sep 28;12(1):5683. doi: 10.1038/s41467-021-25936-2. Nat Commun. 2021. PMID: 34584096 Free PMC article.
-
Suitability of double-stranded DNA as a molecular standard for the validation of analytical ultracentrifugation instruments.Eur Biophys J. 2023 Jul;52(4-5):267-280. doi: 10.1007/s00249-023-01671-y. Epub 2023 Jul 27. Eur Biophys J. 2023. PMID: 37501021 Free PMC article.
-
Supercoiling biases the formation of loops involved in gene regulation.Biophys Rev. 2016 Nov;8(Suppl 1):65-74. doi: 10.1007/s12551-016-0211-0. Epub 2016 Jul 5. Biophys Rev. 2016. PMID: 28510212 Free PMC article. Review.
-
Growth inhibition mediated by excess negative supercoiling: the interplay between transcription elongation, R-loop formation and DNA topology.Mol Microbiol. 2006 Feb;59(3):723-30. doi: 10.1111/j.1365-2958.2005.05006.x. Mol Microbiol. 2006. PMID: 16420346 Review.
References
-
- Franklin R. E. and Gosling R. G. (4, 1953) Molecular configuration in sodium thymonucleate. Nature, 171(4356), 740–741. - PubMed
-
- Lieberman-Aiden E., van Berkum N. L., Williams L., Imakaev M., Ragoczy T., Telling A., Amit I., Lajoie B. R., Sabo P. J., Dorschner M. O., Sandstrom R., Bernstein B., Bender M. A., Groudine M., Gnirke A., Stamatoyannopoulos J., Mirny L. A., Lander E. S., and Dekker J. (10, 2009) Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science (New York, N.Y.), 326(5950), 289–293 - PMC - PubMed
-
- Dorman C. J. (2013) Genome architecture and global generegulation in bacteria: making progress towards a unified model?. Nature Reviews. Microbiology, 11(5), 349–355 - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources