

In Vitro and In Vivo Characterization of H5N8 High-Pathogenicity Avian Influenza Virus Neurotropism in Ducks and Chickens
- PMID: 36625654
- PMCID: PMC9927090
- DOI: 10.1128/spectrum.04229-22
In Vitro and In Vivo Characterization of H5N8 High-Pathogenicity Avian Influenza Virus Neurotropism in Ducks and Chickens
Abstract
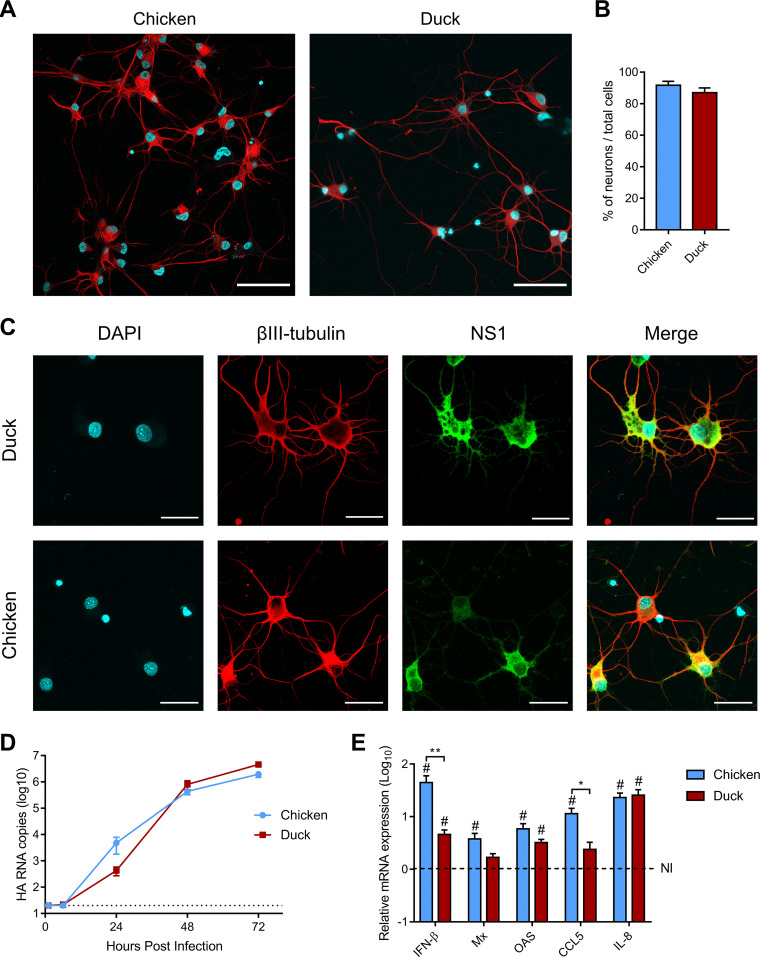
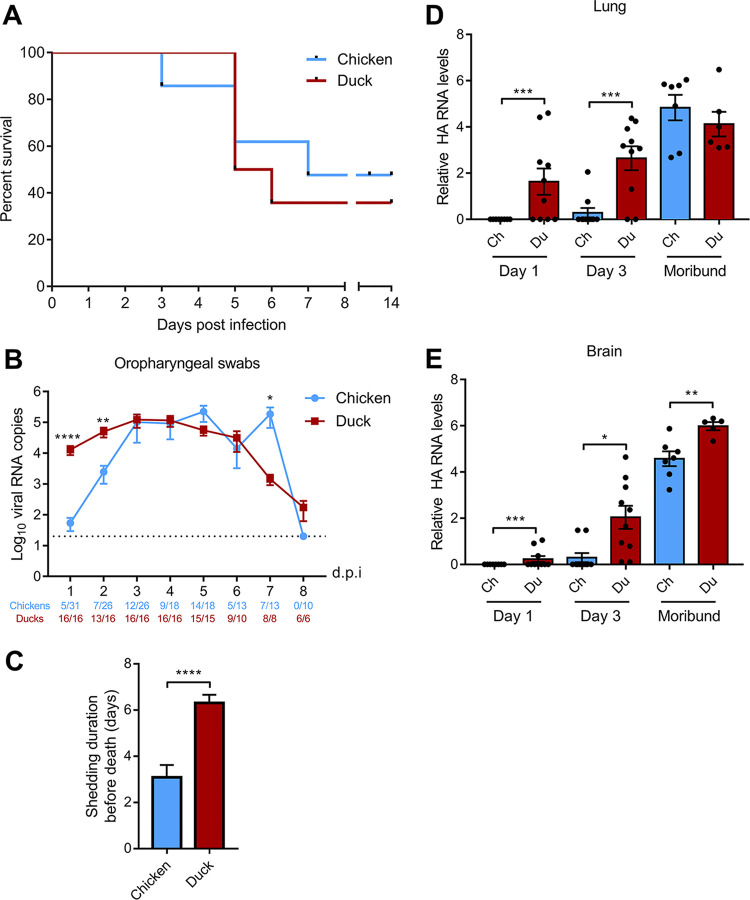
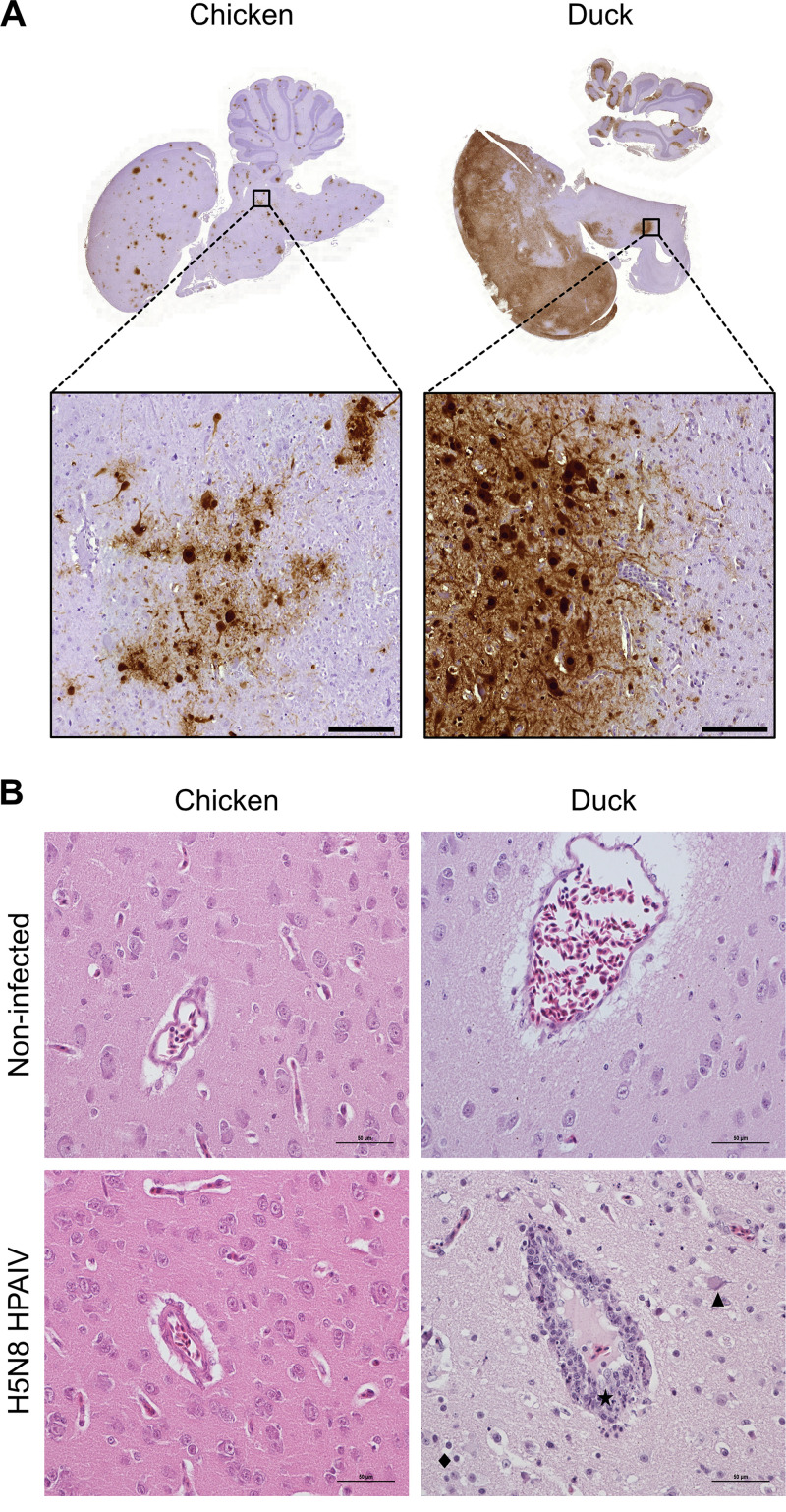
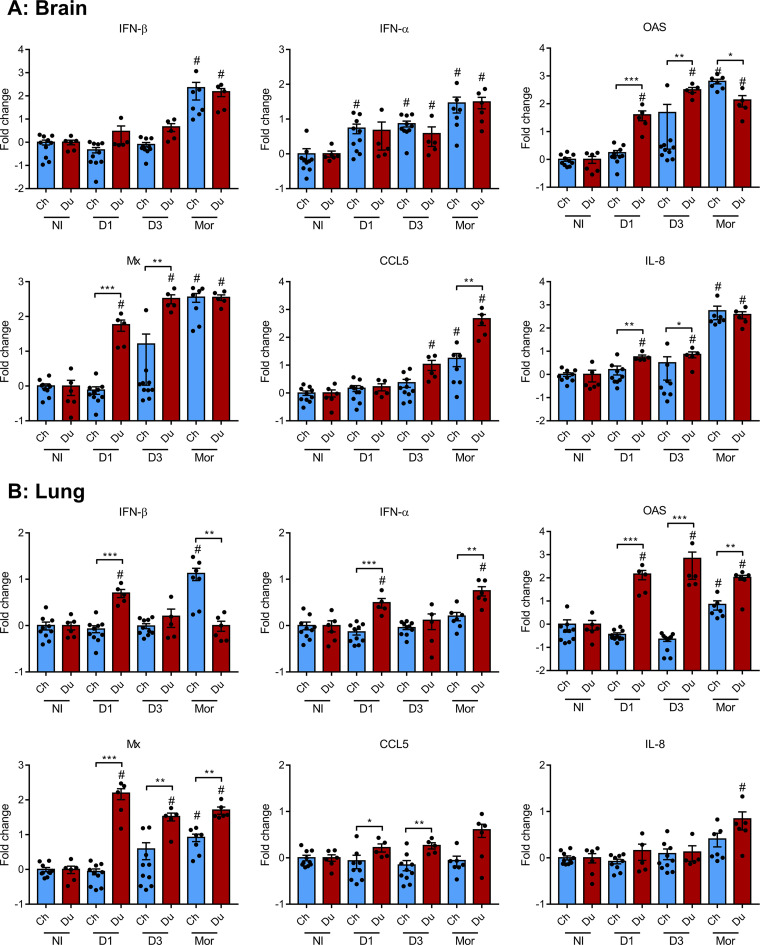
H5N8 high-pathogenicity avian influenza virus (HPAIV) of clade 2.3.4.4B, which circulated during the 2016 epizootics in Europe, was notable for causing different clinical signs in ducks and chickens. The clinical signs preceding death were predominantly neurological in ducks versus respiratory in chickens. To investigate the determinants for the predominant neurological signs observed in ducks, we infected duck and chicken primary cortical neurons. Viral replication was identical in neuronal cultures from both species. In addition, we did not detect any major difference in the immune and inflammatory responses. These results suggest that the predominant neurological involvement of H5N8 HPAIV infection in ducks could not be recapitulated in primary neuronal cultures. In vivo, H5N8 HPAIV replication in ducks peaked soon after infection and led to an early colonization of the central nervous system. In contrast, viral replication was delayed in chickens but ultimately burst in the lungs of chickens, and the chickens died of respiratory distress before brain damage became significant. Consequently, the immune and inflammatory responses in the brain were significantly higher in duck brains than those in chickens. Our study thus suggests that early colonization of the central nervous system associated with prolonged survival after the onset of virus replication is the likely primary cause of the sustained inflammatory response and subsequent neurological disorders observed in H5N8 HPAIV-infected ducks. IMPORTANCE The severity of high-pathogenicity avian influenza virus (HPAIV) infection has been linked to its ability to replicate systemically and cause lesions in a variety of tissues. However, the symptomatology depends on the host species. The H5N8 virus of clade 2.3.4.4B had a pronounced neurotropism in ducks, leading to severe neurological disorders. In contrast, neurological signs were rarely observed in chickens, which suffered mostly from respiratory distress. Here, we investigated the determinants of H5N8 HPAIV neurotropism. We provide evidence that the difference in clinical signs was not due to a difference in neurotropism. Our results rather indicate that chickens died of respiratory distress due to intense viral replication in the lungs before viral replication in the brain could produce significant lesions. In contrast, ducks better controlled virus replication in the lungs, thus allowing the virus to replicate for a sufficient duration in the brain, to reach high levels, and to cause significant lesions.
Keywords: chicken; duck; highly pathogenic avian influenza; influenza; neuron; neurotropism; primary.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Preferential Selection and Contribution of Non-Structural Protein 1 (NS1) to the Efficient Transmission of Panzootic Avian Influenza H5N8 Virus Clades 2.3.4.4A and B in Chickens and Ducks.J Virol. 2021 Aug 25;95(18):e0044521. doi: 10.1128/JVI.00445-21. Epub 2021 Aug 25. J Virol. 2021. PMID: 34160261 Free PMC article.
-
Genetic Determinants for Virulence and Transmission of the Panzootic Avian Influenza Virus H5N8 Clade 2.3.4.4 in Pekin Ducks.J Virol. 2022 Jul 13;96(13):e0014922. doi: 10.1128/jvi.00149-22. Epub 2022 Jun 7. J Virol. 2022. PMID: 35670594 Free PMC article.
-
Opposite Outcomes of the Within-Host Competition between High- and Low-Pathogenic H5N8 Avian Influenza Viruses in Chickens Compared to Ducks.J Virol. 2022 Jan 12;96(1):e0136621. doi: 10.1128/JVI.01366-21. Epub 2021 Oct 6. J Virol. 2022. PMID: 34613804 Free PMC article.
-
Evaluation of three hemagglutinin-based vaccines for the experimental control of a panzootic clade 2.3.4.4b A(H5N8) high pathogenicity avian influenza virus in mule ducks.Vaccine. 2023 Jan 4;41(1):145-158. doi: 10.1016/j.vaccine.2022.11.012. Epub 2022 Nov 18. Vaccine. 2023. PMID: 36411134
-
The genetics of highly pathogenic avian influenza viruses of subtype H5 in Germany, 2006-2020.Transbound Emerg Dis. 2021 May;68(3):1136-1150. doi: 10.1111/tbed.13843. Epub 2020 Sep 29. Transbound Emerg Dis. 2021. PMID: 32964686 Review.
Cited by
-
High pathogenicity avian influenza A (H5N1) clade 2.3.4.4b virus infection in a captive Tibetan black bear (Ursus thibetanus): investigations based on paraffin-embedded tissues, France, 2022.Microbiol Spectr. 2024 Mar 5;12(3):e0373623. doi: 10.1128/spectrum.03736-23. Epub 2024 Feb 2. Microbiol Spectr. 2024. PMID: 38305177 Free PMC article.
-
Highly Pathogenic Avian Influenza (HPAI) H5 Clade 2.3.4.4b Virus Infection in Birds and Mammals.Animals (Basel). 2024 May 2;14(9):1372. doi: 10.3390/ani14091372. Animals (Basel). 2024. PMID: 38731377 Free PMC article. Review.
-
Mass mortality event in South American sea lions (Otaria flavescens) correlated to highly pathogenic avian influenza (HPAI) H5N1 outbreak in Chile.Vet Q. 2023 Dec;43(1):1-10. doi: 10.1080/01652176.2023.2265173. Epub 2023 Oct 19. Vet Q. 2023. PMID: 37768676 Free PMC article.
References
-
- Pohlmann A, King J, Fusaro A, Zecchin B, Banyard AC, Brown IH, Byrne AMP, Beerens N, Liang Y, Heutink R, Harders F, James J, Reid SM, Hansen RDE, Lewis NS, Hjulsager C, Larsen LE, Zohari S, Anderson K, Bröjer C, Nagy A, Savič V, van Borm S, Steensels M, Briand F-X, Swieton E, Smietanka K, Grund C, Beer M, Harder T. 2022. Has epizootic become enzootic? Evidence for a fundamental change in the infection dynamics of highly pathogenic avian influenza in Europe, 2021. mBio 13:e0060922. doi:10.1128/mbio.00609-22. - DOI - PMC - PubMed
-
- Pyankova OG, Susloparov IM, Moiseeva AA, Kolosova NP, Onkhonova GS, Danilenko AV, Vakalova EV, Shendo GL, Nekeshina NN, Noskova LN, Demina JV, Frolova NV, Gavrilova EV, Maksyutov RA, Ryzhikov AB. 2021. Isolation of clade 2.3.4.4b A(H5N8), a highly pathogenic avian influenza virus, from a worker during an outbreak on a poultry farm, Russia, December 2020. Eurosurveillance 26:2100439. doi:10.2807/1560-7917.ES.2021.26.24.2100439. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
