

Quantification of substoichiometric modification reveals global tsRNA hypomodification, preferences for angiogenin-mediated tRNA cleavage, and idiosyncratic epitranscriptomes of human neuronal cell-lines
- PMID: 36618980
- PMCID: PMC9798144
- DOI: 10.1016/j.csbj.2022.12.020
Quantification of substoichiometric modification reveals global tsRNA hypomodification, preferences for angiogenin-mediated tRNA cleavage, and idiosyncratic epitranscriptomes of human neuronal cell-lines
Abstract
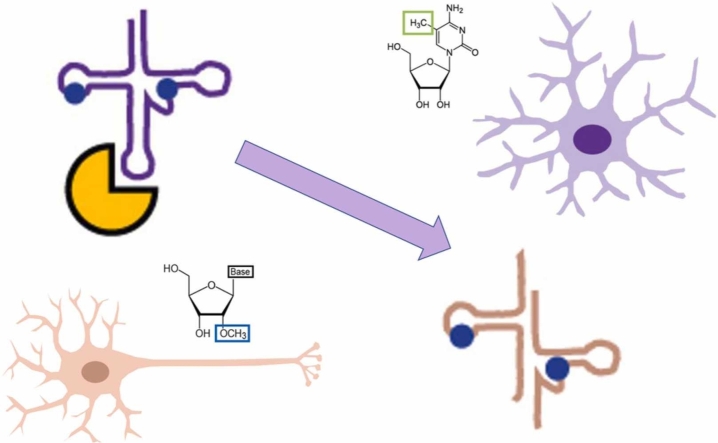
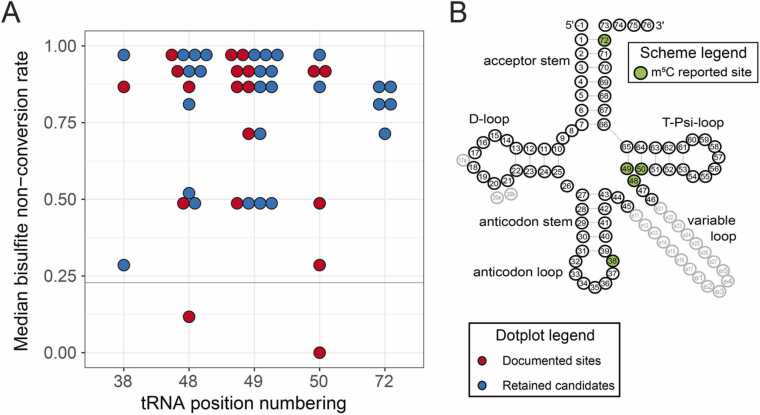
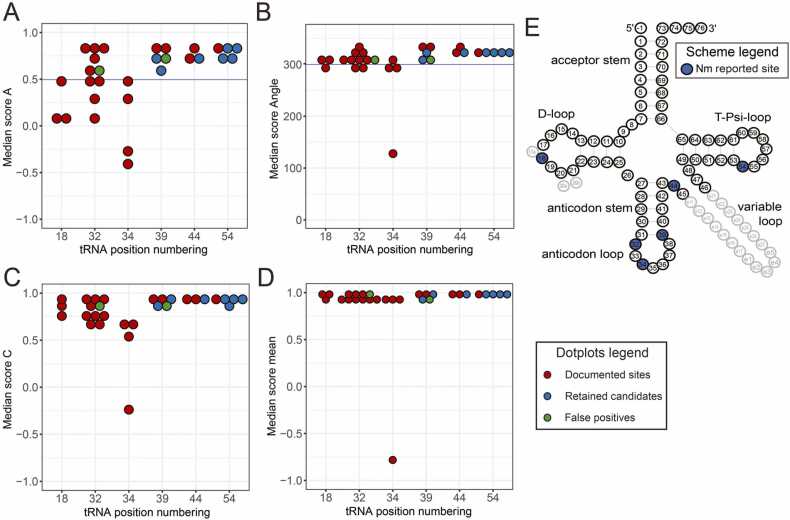
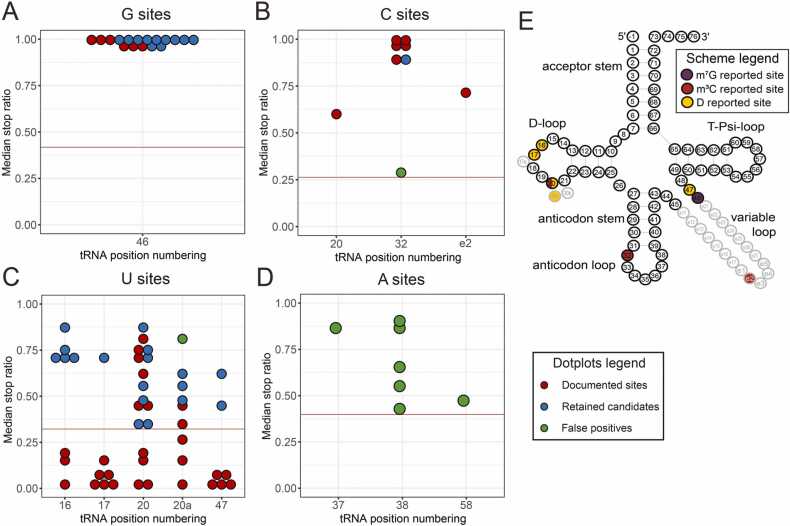
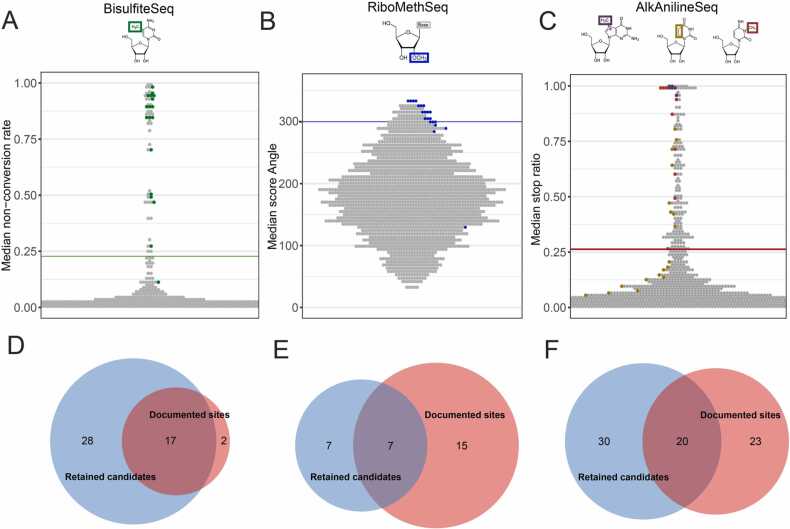
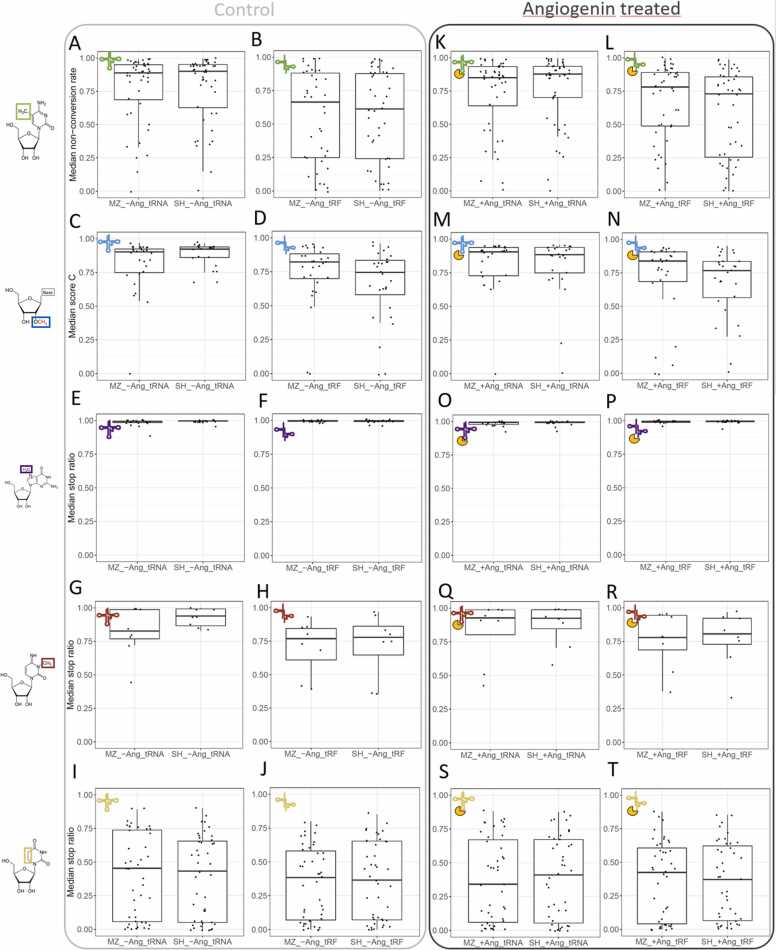
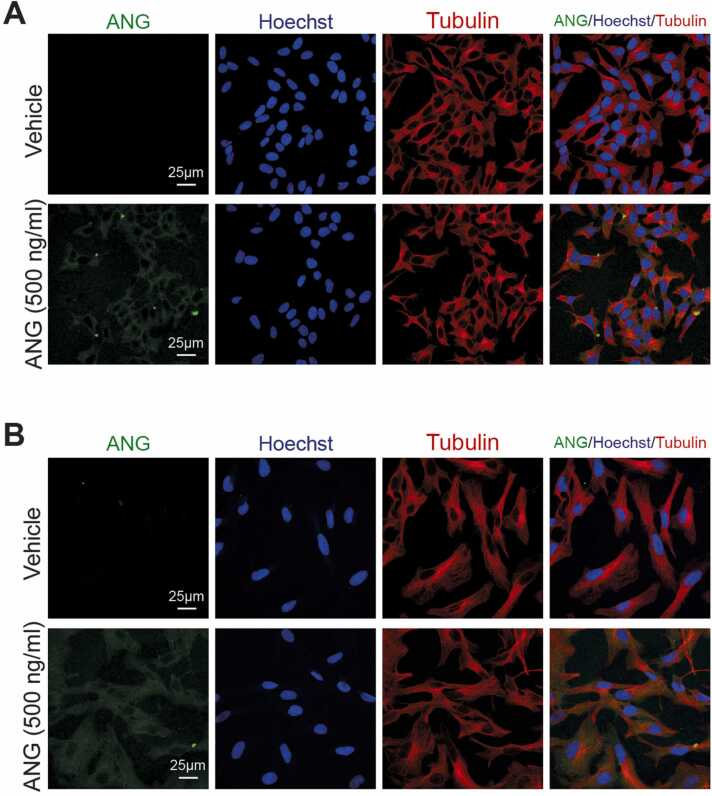
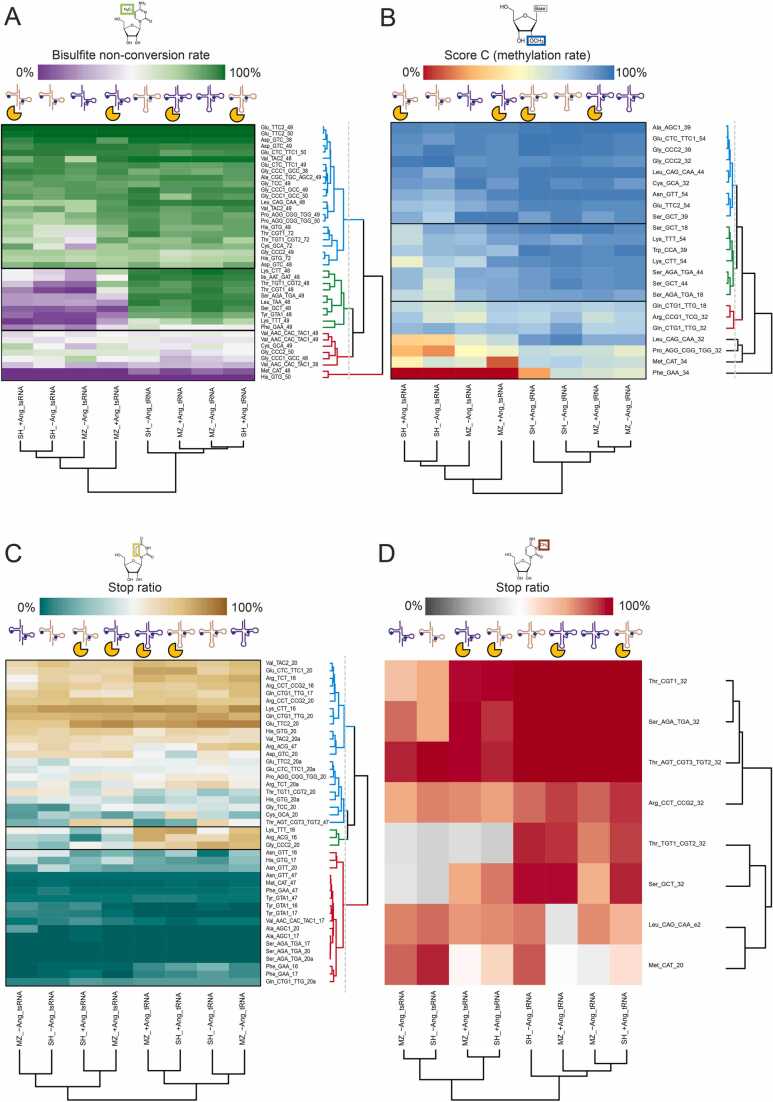
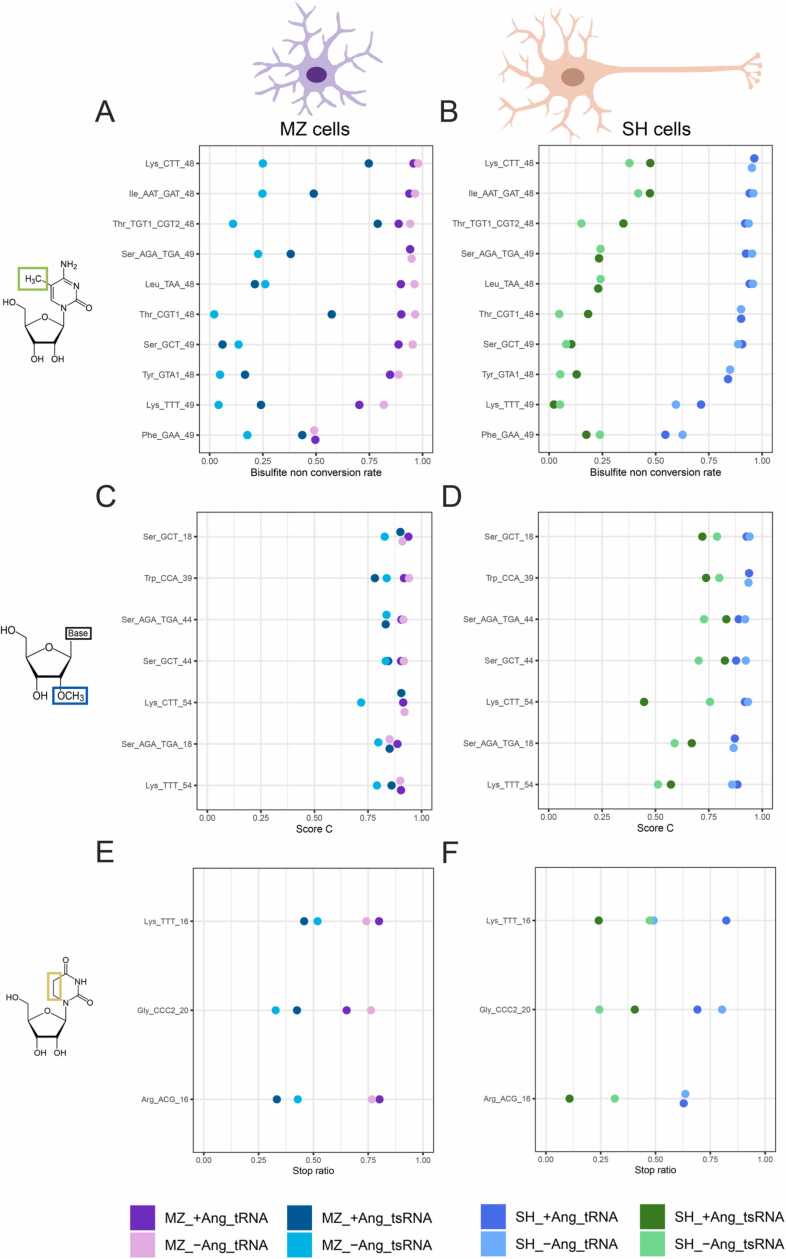
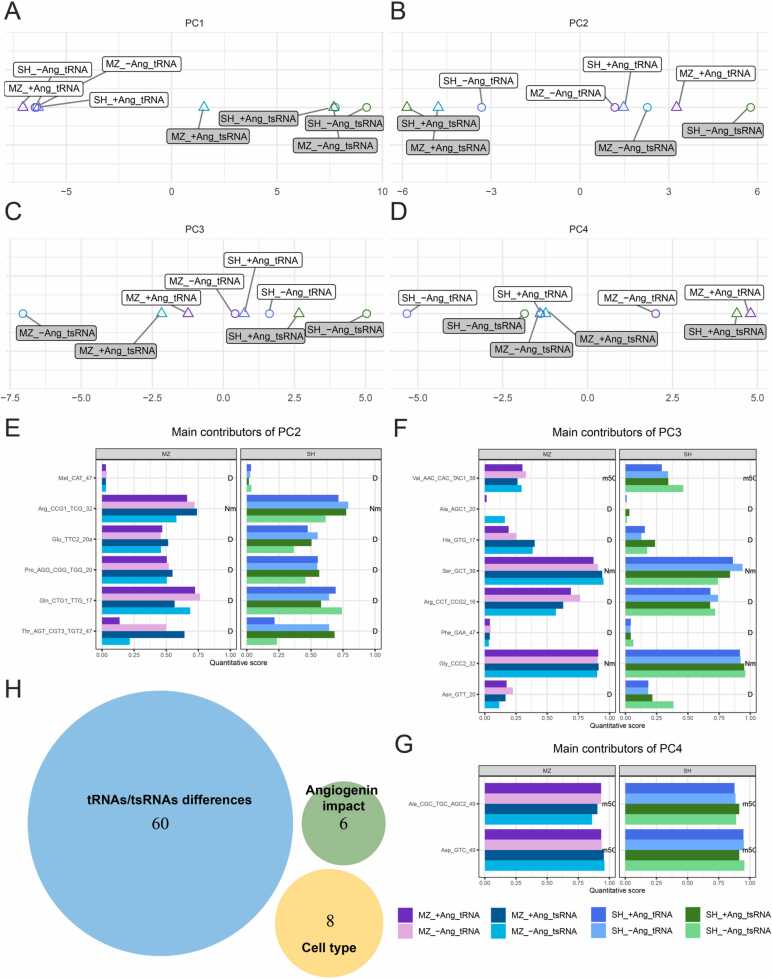
Modification of tRNA is an integral part of the epitranscriptome with a particularly pronounced potential to generate diversity in RNA expression. Eukaryotic tRNA contains modifications in up to 20% of their nucleotides, but not all sites are always fully modified. Combinations and permutations of partially modified sites in tRNAs can generate a plethora of tRNA isoforms, termed modivariants. Here, we investigate the stoichiometry of incompletely modified sites in tRNAs from human cell lines for their information content. Using a panel of RNA modification mapping methods, we assess the stoichiometry of sites that contain the modifications 5-methylcytidine (m5C), 2'-O-ribose methylation (Nm), 3-methylcytidine (m3C), 7-methylguanosine (m7G), and Dihydrouridine (D). We discovered that up to 75% of sites can be incompletely modified and that the differential modification status of a cellular tRNA population holds information that allows to discriminate e.g. different cell lines. As a further aspect, we investigated potential causal connectivity between tRNA modification and its processing into tRNA fragments (tiRNAs and tRFs). Upon exposure of cultured living cells to cell-penetrating angiogenin, the modification patterns of the corresponding RNA populations was changed. Importantly, we also found that tsRNAs were significantly less modified than their parent tRNAs at numerous sites, suggesting that tsRNAs might derive chiefly from hypomodified tRNAs.
Keywords: Angiogenin; Modification; Modification mapping; RNAseq; TRNA; TRNA fragments.
© 2022 The Author(s).
Conflict of interest statement
Mark Helm is a consultant for Moderna Inc.
Figures
Similar articles
-
m5U54 tRNA Hypomodification by Lack of TRMT2A Drives the Generation of tRNA-Derived Small RNAs.Int J Mol Sci. 2021 Mar 14;22(6):2941. doi: 10.3390/ijms22062941. Int J Mol Sci. 2021. PMID: 33799331 Free PMC article.
-
Genome-wide analysis of N1-methyl-adenosine modification in human tRNAs.RNA. 2010 Jul;16(7):1317-27. doi: 10.1261/rna.2057810. Epub 2010 May 19. RNA. 2010. PMID: 20484468 Free PMC article.
-
tsRNA modifications: An emerging layer of biological regulation in disease.J Adv Res. 2024 Sep 10:S2090-1232(24)00401-6. doi: 10.1016/j.jare.2024.09.010. Online ahead of print. J Adv Res. 2024. PMID: 39260796 Review.
-
Queuosine modification protects cognate tRNAs against ribonuclease cleavage.RNA. 2018 Oct;24(10):1305-1313. doi: 10.1261/rna.067033.118. Epub 2018 Jul 3. RNA. 2018. PMID: 29970597 Free PMC article.
-
Transfer RNA-derived fragments and tRNA halves: biogenesis, biological functions and their roles in diseases.J Mol Med (Berl). 2018 Nov;96(11):1167-1176. doi: 10.1007/s00109-018-1693-y. Epub 2018 Sep 19. J Mol Med (Berl). 2018. PMID: 30232504 Review.
Cited by
-
Biological function and clinical application prospect of tsRNAs in digestive system biology and pathology.Cell Commun Signal. 2023 Oct 30;21(1):302. doi: 10.1186/s12964-023-01341-8. Cell Commun Signal. 2023. PMID: 37904174 Free PMC article. Review.
-
Methylation modifications in tRNA and associated disorders: Current research and potential therapeutic targets.Cell Prolif. 2024 Sep;57(9):e13692. doi: 10.1111/cpr.13692. Epub 2024 Jun 28. Cell Prolif. 2024. PMID: 38943267 Free PMC article. Review.
-
Translational response to mitochondrial stresses is orchestrated by tRNA modifications.bioRxiv [Preprint]. 2024 Feb 14:2024.02.14.580389. doi: 10.1101/2024.02.14.580389. bioRxiv. 2024. PMID: 38405984 Free PMC article. Preprint.
-
General Principles and Limitations for Detection of RNA Modifications by Sequencing.Acc Chem Res. 2024 Feb 6;57(3):275-288. doi: 10.1021/acs.accounts.3c00529. Epub 2023 Dec 8. Acc Chem Res. 2024. PMID: 38065564 Free PMC article.
-
tRNA-derived fragments: mechanism of gene regulation and clinical application in lung cancer.Cell Oncol (Dordr). 2024 Feb;47(1):37-54. doi: 10.1007/s13402-023-00864-z. Epub 2023 Aug 29. Cell Oncol (Dordr). 2024. PMID: 37642916 Review.
References
-
- Rashad S., Tominaga T., Niizuma K. The cell and stress-specific canonical and noncanonical tRNA cleavage. J Cell Physiol. 2021;236(5):3710–3724. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials