

Ganoin and acrodin formation on scales and teeth in spotted gar: A vital role of enamelin in the unique process of enamel mineralization
- PMID: 36464775
- PMCID: PMC10239528
- DOI: 10.1002/jez.b.23183
Ganoin and acrodin formation on scales and teeth in spotted gar: A vital role of enamelin in the unique process of enamel mineralization
Abstract
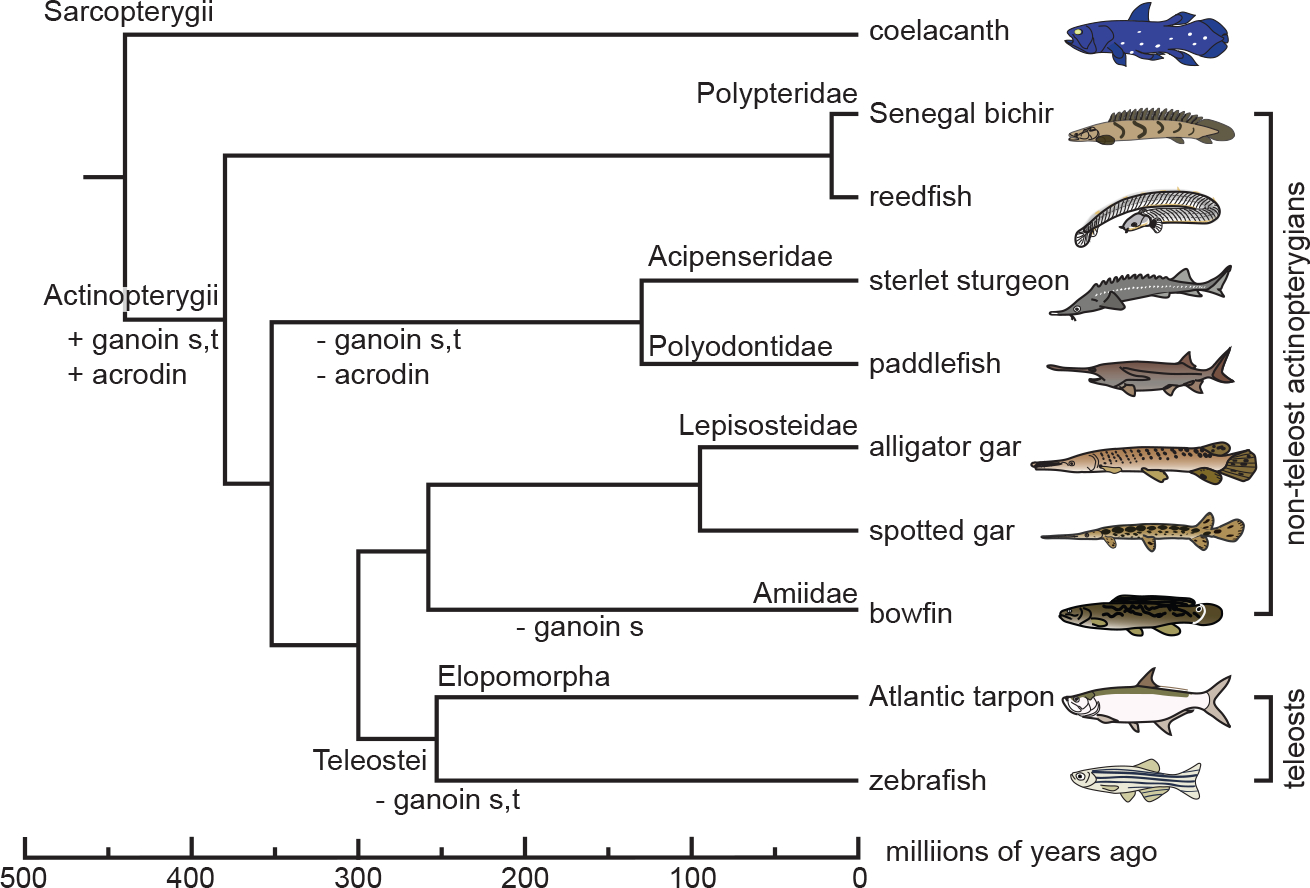
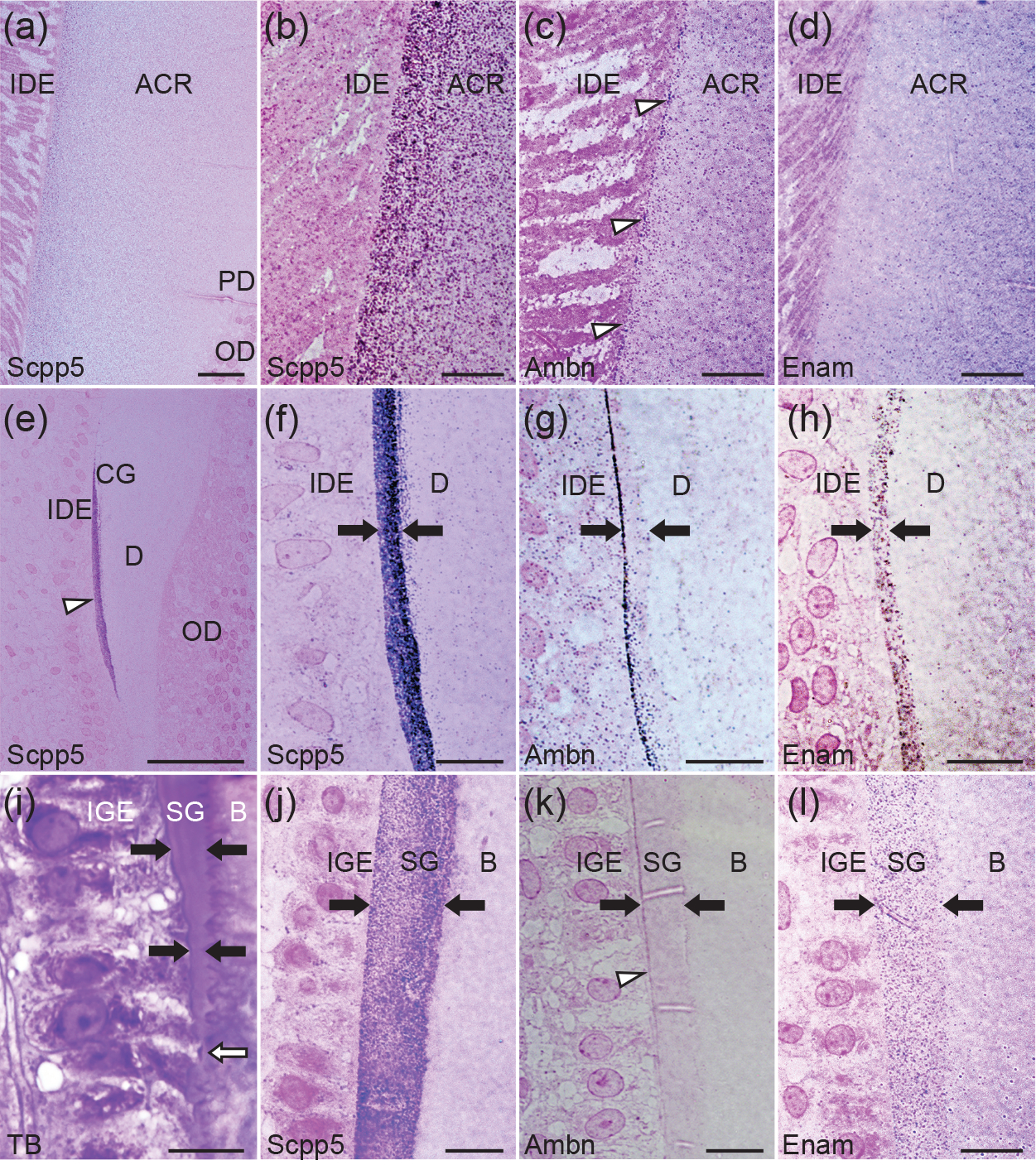
Gars and bichirs develop scales and teeth with ancient actinopterygian characteristics. Their scale surface and tooth collar are covered with enamel, also known as ganoin, whereas the tooth cap is equipped with an enamel-like tissue, acrodin. Here, we investigated the formation and mineralization of the ganoin and acrodin matrices in spotted gar, and the evolution of the scpp5, ameloblastin (ambn), and enamelin (enam) genes, which encode matrix proteins of ganoin. Results suggest that, in bichirs and gars, all these genes retain structural characteristics of their orthologs in stem actinopterygians, presumably reflecting the presence of ganoin on scales and teeth. During scale formation, Scpp5 and Enam were initially found in the incipient ganoin matrix and the underlying collagen matrix, whereas Ambn was detected mostly in a surface region of the well-developed ganoin matrix. Although collagen is the principal acrodin matrix protein, Scpp5 was detected within the matrix. Similarities in timings of mineralization and the secretion of Scpp5 suggest that acrodin evolved by the loss of the matrix secretory stage of ganoin formation: dentin formation is immediately followed by the maturation stage. The late onset of Ambn secretion during ganoin formation implies that Ambn is not essential for mineral ribbon formation, the hallmark of the enamel matrix. Furthermore, Scpp5 resembles amelogenin that is not important for the initial formation of mineral ribbons in mammals. It is thus likely that the evolution of ENAM was vital to the origin of the unique mineralization process of the enamel matrix.
Keywords: SCPP genes; SCPP5; ameloblastin; enamelin; ganoid scales; mineral ribbons.
© 2022 Wiley Periodicals LLC.
Conflict of interest statement
Declaration of Competing Interest: The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures



Similar articles
-
Coevolution of enamel, ganoin, enameloid, and their matrix SCPP genes in osteichthyans.iScience. 2021 Jan 1;24(1):102023. doi: 10.1016/j.isci.2020.102023. eCollection 2021 Jan 22. iScience. 2021. PMID: 33506188 Free PMC article.
-
New genomic and fossil data illuminate the origin of enamel.Nature. 2015 Oct 1;526(7571):108-11. doi: 10.1038/nature15259. Epub 2015 Sep 23. Nature. 2015. PMID: 26416752
-
Immunolocalization of enamel matrix protein-like proteins in the tooth enameloid of spotted gar, Lepisosteus oculatus, an actinopterygian bony fish.Connect Tissue Res. 2019 May;60(3):291-303. doi: 10.1080/03008207.2018.1506446. Epub 2018 Aug 13. Connect Tissue Res. 2019. PMID: 30063414
-
The developing enamel matrix: nature and function.Eur J Oral Sci. 1998 Jan;106 Suppl 1:282-91. doi: 10.1111/j.1600-0722.1998.tb02188.x. Eur J Oral Sci. 1998. PMID: 9541238 Review.
-
Enamelin and autosomal-dominant amelogenesis imperfecta.Crit Rev Oral Biol Med. 2003;14(6):387-98. doi: 10.1177/154411130301400602. Crit Rev Oral Biol Med. 2003. PMID: 14656895 Review.
Cited by
-
The spotted parrotfish genome provides insights into the evolution of a coral reef dietary specialist (Teleostei: Labridae: Scarini: Cetoscarus ocellatus).Ecol Evol. 2024 Mar 12;14(3):e11148. doi: 10.1002/ece3.11148. eCollection 2024 Mar. Ecol Evol. 2024. PMID: 38476702 Free PMC article.
References
-
- Al-Hashimi N, Sire J-Y, & Delgado S (2009). Evolutionary analysis of mammalian enamelin, the largest enamel protein, supports a crucial role for the 32-kDa peptide and reveals selective adaptation in rodents and primates. Journal of Molecular Evolution 69:635–656. 10.1007/s00239-009-9302-x - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
