

Structural and biochemical basis of interdependent FANCI-FANCD2 ubiquitination
- PMID: 36385258
- PMCID: PMC9890228
- DOI: 10.15252/embj.2022111898
Structural and biochemical basis of interdependent FANCI-FANCD2 ubiquitination
Abstract
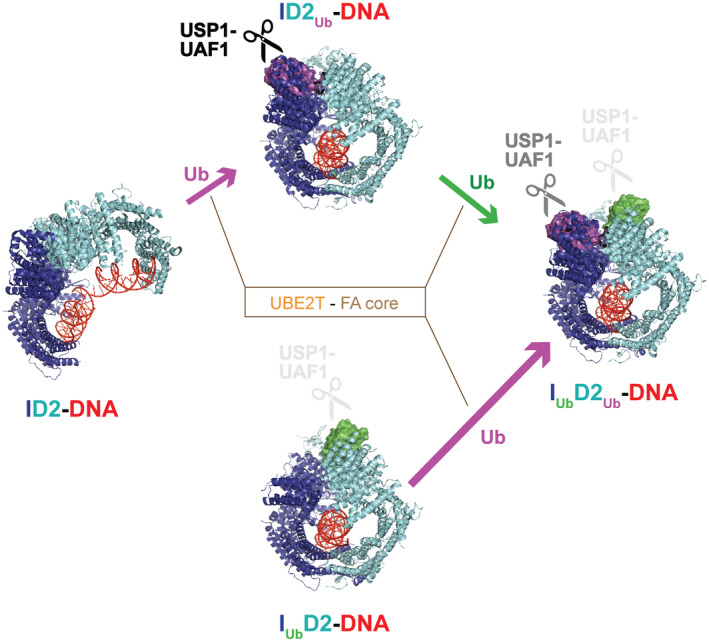
Di-monoubiquitination of the FANCI-FANCD2 (ID2) complex is a central and crucial step for the repair of DNA interstrand crosslinks via the Fanconi anaemia pathway. While FANCD2 ubiquitination precedes FANCI ubiquitination, FANCD2 is also deubiquitinated at a faster rate than FANCI, which can result in a FANCI-ubiquitinated ID2 complex (IUb D2). Here, we present a 4.1 Å cryo-EM structure of IUb D2 complex bound to double-stranded DNA. We show that this complex, like ID2Ub and IUb D2Ub , is also in the closed ID2 conformation and clamps on DNA. The target lysine of FANCD2 (K561) becomes fully exposed in the IUb D2-DNA structure and is thus primed for ubiquitination. Similarly, FANCI's target lysine (K523) is also primed for ubiquitination in the ID2Ub -DNA complex. The IUb D2-DNA complex exhibits deubiquitination resistance, conferred by the presence of DNA and FANCD2. ID2Ub -DNA, on the other hand, can be efficiently deubiquitinated by USP1-UAF1, unless further ubiquitination on FANCI occurs. Therefore, FANCI ubiquitination effectively maintains FANCD2 ubiquitination in two ways: it prevents excessive FANCD2 deubiquitination within an IUb D2Ub -DNA complex, and it enables re-ubiquitination of FANCD2 within a transient, closed-on-DNA, IUb D2 complex.
Keywords: DNA binding; Fanconi anaemia; USP1-UAF1; deubiquitination; interstrand crosslink repair.
© 2022 The Authors. Published under the terms of the CC BY 4.0 license.
Figures
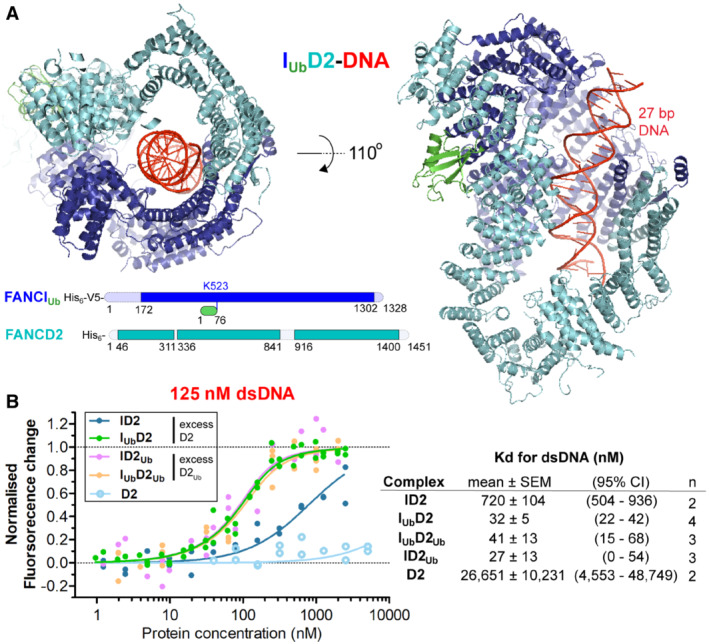
FANCIUb‐FANCD2 (IUbD2) structure bound to double‐stranded DNA. The structure was determined by cryo‐EM, using a 4.1 Å global resolution map. Two different views of the structure are shown. Unmodelled regions (due to poor density) extending 20 amino‐acid stretches are indicated at the bottom.
Ubiquitination of either subunit of the ID2 complex results in increased affinity to double‐stranded DNA (dsDNA). Left: Normalised fluorescent changes of IRDye700‐labelled 32 bp DNA (125 nM) when incubated with increasing concentrations (ranging from 1 nM to 2.5 μM) of FANCI (I) or ubiquitinated FANCI (IUb) in the presence of a constant excess concentration (equal to the maximum concentration of I/IUb used) of FANCD2 (D2) or ubiquitinated FANCD2 (D2Ub). As a control, normalised fluorescent changes of IRDye700‐labelled DNA (125 nM) when incubated with increasing concentrations of FANCD2 (ranging from 40 nM to 5 μM) were monitored as well. For each protein/complex, the experiment was conducted 2–4 times (technical replicates) and all data points from the replicate experiments were used for fitting of a one‐site binding model. Right: Apparent ID2, IUbD2 and D2 Kd values (and associated uncertainties, all in nM) for dsDNA measured from model fitting (n: number of binding experiments per protein/complex).
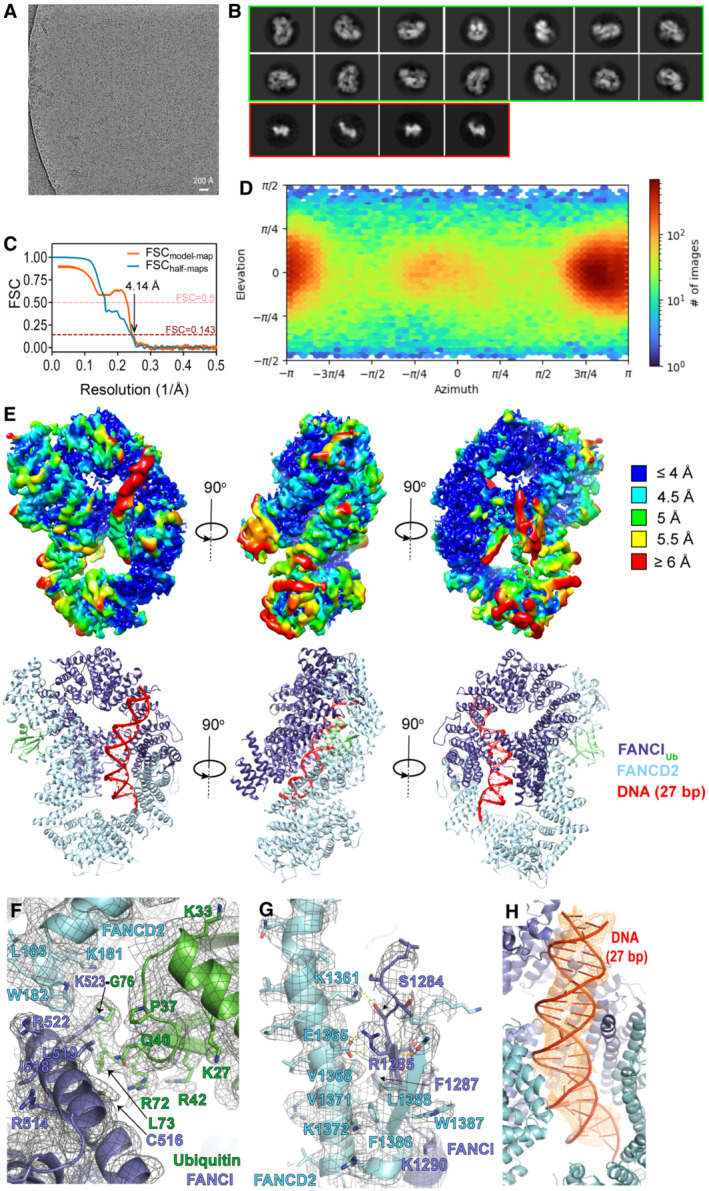
Example micrograph with scale bar.
Example 2D classes. Circular mask is 170 Å in diameter. 2D classes surrounded by a green box correspond to IUbD2‐DNA complex particles, while smaller‐sized 2D classes surrounded by a red box likely correspond to monomeric IUb/D2 proteins.
Fourier Shell Correlation (FSC) curves: between the two half maps produced in the final local non‐uniform refinement (shown in blue) and between the refined model and final map (shown in orange).
Particle orientation (viewing direction distribution) in the final map. Total number of particles: 139,601.
Top: Locally filtered map coloured by local resolution, viewed from three different angles. Bottom: Corresponding structural model viewed under same orientations.
IUbD2‐DNA structure with corresponding map density (locally filtered map), centred on the isopeptide bond between K523 of FANCI and G76 of ubiquitin. Some well‐resolved side‐chains are illustrated as sticks and indicated.
Interaction between FANCI and FANCD2 C‐termini with corresponding map density (locally filtered map). A beta‐sheet consisting of a FANCI and a FANCD2 strand is formed (residues 1,285–1,289 of FANCI and residues 1,384–1,388 of FANCD2). This is held in place through hydrophobic and electrostatic interactions with a FANCD2 helix (1,351–1,377 aa). Residues predicted to participate in such interactions are shown as sticks and indicated. Selected side chains, for which there is good density are also shown as sticks. For clarity, adjacent to that region elements of the IUbD2‐DNA structure and map are not shown.
IUbD2‐DNA structure centred on DNA. Density corresponding to the 27 bp modelled DNA is shown as orange mesh. Colouring of structure is as in (E–G).
The absence of a FANCD2‐conjugated ubiquitin in IUbD2‐DNA structure is associated with a disorder in the N‐terminus of FANCI (residues 1–170), when compared with IUbD2Ub‐DNA structure. The two structures were aligned in Pymol and visualised from the same angle, either on their own (left and centre), or together (right).
Helices of FANCI involved in interaction with FANCD2's ubiquitin (IUbD2Ub‐DNA structure; FANCI: blue, Ubiquitin: magenta), are positioned differently when the ubiquitin is removed (IUbD2‐DNA structure; FANCI: orange). IUbD2Ub‐DNA and IUbD2‐DNA structures were fitted to IUbD2‐DNA sharpened map (yellow) and centred on FANCI N‐terminus.
Removal of ubiquitin (magenta) from FANCD2 results in slight movements affecting several FANCD2 helices, from α31 (helix where ubiquitin is conjugated) up to α18. FANCD2 helices of IUbD2‐DNA and IUbD2Ub‐DNA are better aligned towards the N‐terminus of FANCD2 (N‐terminally to, and including α18 helix of FANCD2), whereby FANCD2 interacts with the ubiquitin (green) conjugated to FANCI. The structures shown in (A), were centred on the central part of FANCD2.
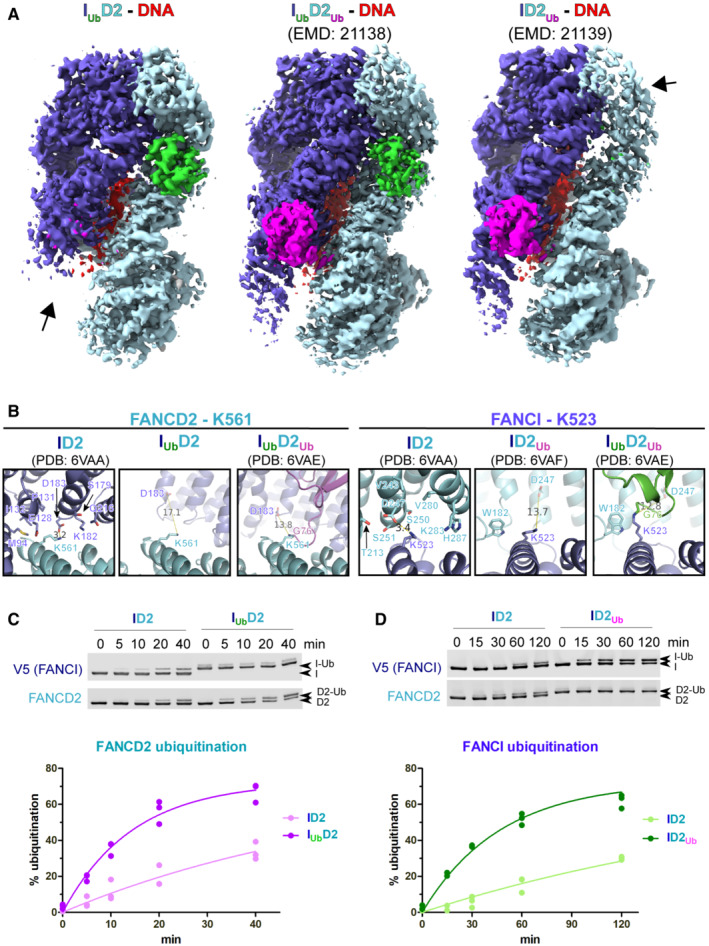
- A
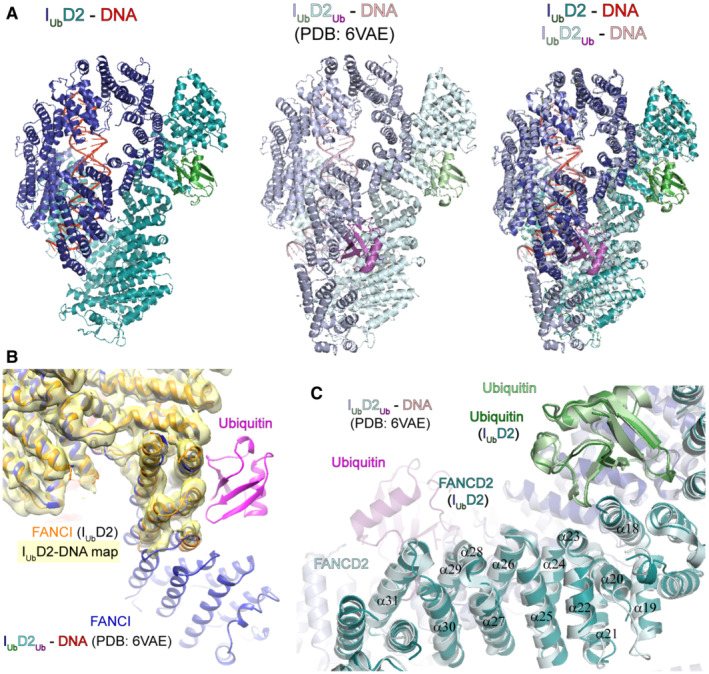
Comparison of cryo‐EM density distribution among IUbD2‐DNA (Phenix‐auto‐sharpened map), IUbD2Ub‐DNA (EMD‐21138) and ID2Ub‐DNA (EMD‐21138) maps. IUbD2‐DNA and ID2Ub‐DNA maps, as well as IUbD2Ub‐DNA model (PDB:
6VAE ) were aligned to IUbD2Ub‐DNA in ChimeraX. A different colour was applied for each of the protein chains of IUbD2Ub‐DNA model (FANsCI: slate blue, Ubiquitin‐on‐FANCI: green, FANCD2: cyan, Ubiquitin‐on‐FANCD2: magenta), while DNA was coloured red. Then each map was coloured according to nearby (within 6 Å) residue colours. Contour levels were adjusted (IUbD2‐DNA: 6.21, IUbD2Ub‐DNA: 0.0194 and ID2Ub‐DNA: 0.0162) to achieve comparable volumes among all displayed maps (ranging from 8.6 to 9.4 × 104 Å3). Arrows indicate regions of poorer density (in IUbD2‐DNA and ID2Ub‐DNA maps) relative to other regions of the map, as well as to equivalent positions in the other two maps. - B
Both K561 of FANCD2 and K523 of FANCI become more accessible upon ubiquitination of the other ID2 subunit. Structural comparison of relative accessibility of FANCD2‐K561, upon FANCI ubiquitination (left panel), and of FANCI‐K523, upon FANCD2 ubiquitination (right panel). The relative positions of these lysines upon conjugation with ubiquitin, are also shown for comparison. Residues of the other ID2 subunit within 8 Å distance from the epsilon‐amino‐group of the corresponding lysine are indicated as sticks. The distance to the nearest residue is shown prior and upon ubiquitination of the other ID2 subunit. In either case this increases, upon ubiquitination of the other subunit, further than 10 Å.
- C, D
ID2 ubiquitination on FANCI results in increased rate of FANCD2 ubiquitination (B), whereas ID2 ubiquitination on FANCD2 results in increased rate of FANCI ubiquitination (C). Protein complexes were assembled in vitro on ice in the presence of dsDNA (32 bp) and their in vitro ubiquitination at 30°C was subsequently monitored in a time‐course: at indicative time‐points, aliquots of the reaction were removed and analysed by western blotting using FANCD2 and V5 antibodies (Top). For each protein complex, data‐points from three replicate experiments (three technical replicates) were used in fitting to a one‐phase association model (Bottom).
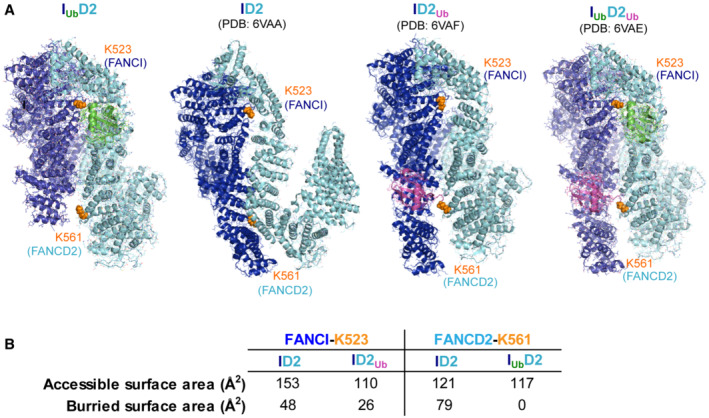
The overall accessibility of FANCI's K523 and FANCD2's K561 is shown within IUbD2‐DNA, ID2‐DNA (PDB:
6VAA ), ID2Ub‐DNA (PDB:6VAF ) and IUbD2Ub‐DNA (PDB:6VAE ) structures. For clarity DNA was removed from the structures. The corresponding lysines are illustrated as orange spheres. IUbD2, ID2Ub and IUbD2Ub structures were aligned to FANCI of ID2 structure, to allow visualisation of all structures under same orientation.FANCI's K523 and FANCD2's K561 accessible surface areas and buried surface area (both in Å2) are shown in non‐ubiquitinated ID2 complex and upon ubiquitination of the other subunit. These values were determined from associated PDB files using the PDBePISA tool (Krissinel & Henrick,
2007 ) athttps://www.ebi.ac.uk/pdbe/pisa/ .
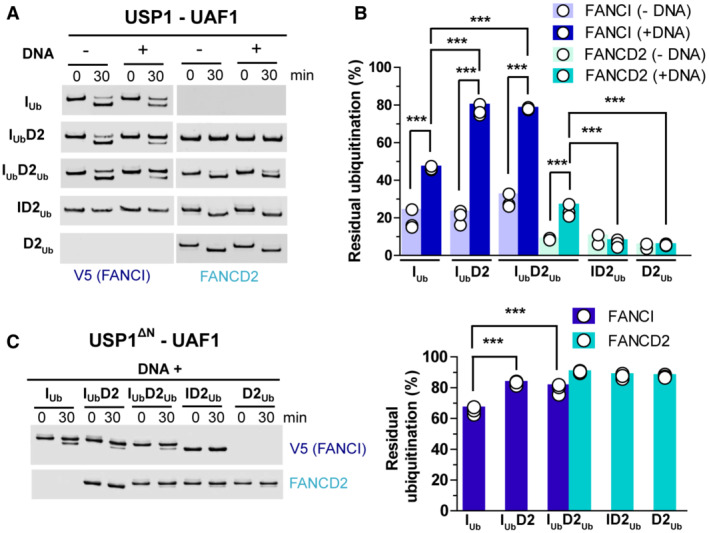
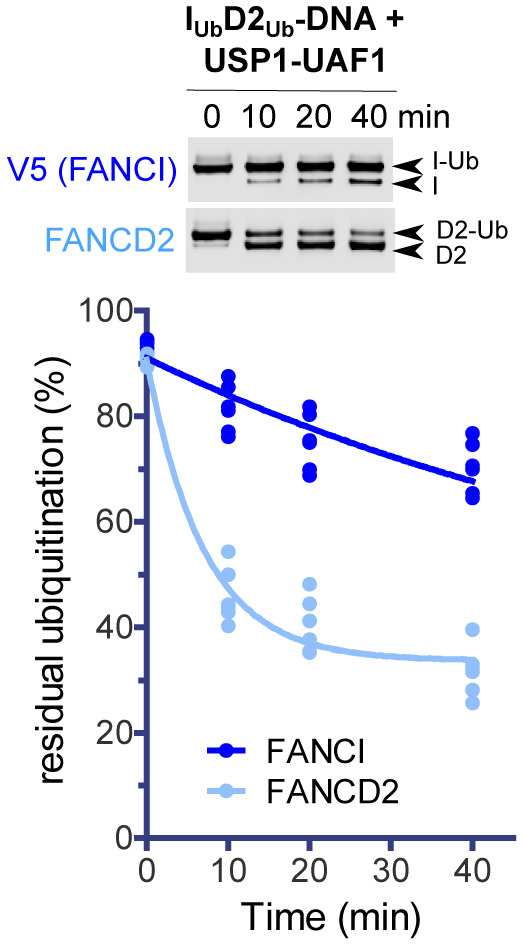
USP1‐UAF1‐mediated deubiquitination of V5‐FANCI and FANCD2 was assessed in the absence or presence of DNA (51 bp), when ubiquitinated versions of these proteins were in isolation (IUb and D2Ub) or within singly/doubly ubiquitinated ID2 complexes (IUbD2, IUbD2Ub and ID2Ub). At indicated time‐points, aliquots of each reaction were removed and analysed by western blotting using FANCD2 and V5 antibodies.
Residual FANCI and FANCD2 ubiquitination following USP1‐UAF1 treatment for 30 min at room temperature. Experiments shown in (A) were performed in triplicate (technical replicates), apart from ID2Ub and D2Ub deubiquitination in the absence of DNA, which were performed twice (and were thus excluded from statistical analysis). Replicate residual ubiquitination values and statistically significant changes (one‐way ANOVA test with Bonferroni correction) are shown. ***P < 0.001.
Deletion of N‐terminus (∆N) of USP1 (residues 1–54) results in greatly reduced FANCD2 deubiquitination. Assays were performed in triplicate (technical replicates) as in (A), but all reactions contained DNA. Left: Western blotting of reaction products at zero and 30 min using FANCD2 and V5 antibodies. Right: Residual FANCI and FANCD2 ubiquitination following USP1‐UAF1 treatment for 30 min. Replicate residual ubiquitination values and statistically significant changes (one‐way ANOVA test with Bonferroni correction) are shown. ***P < 0.001.
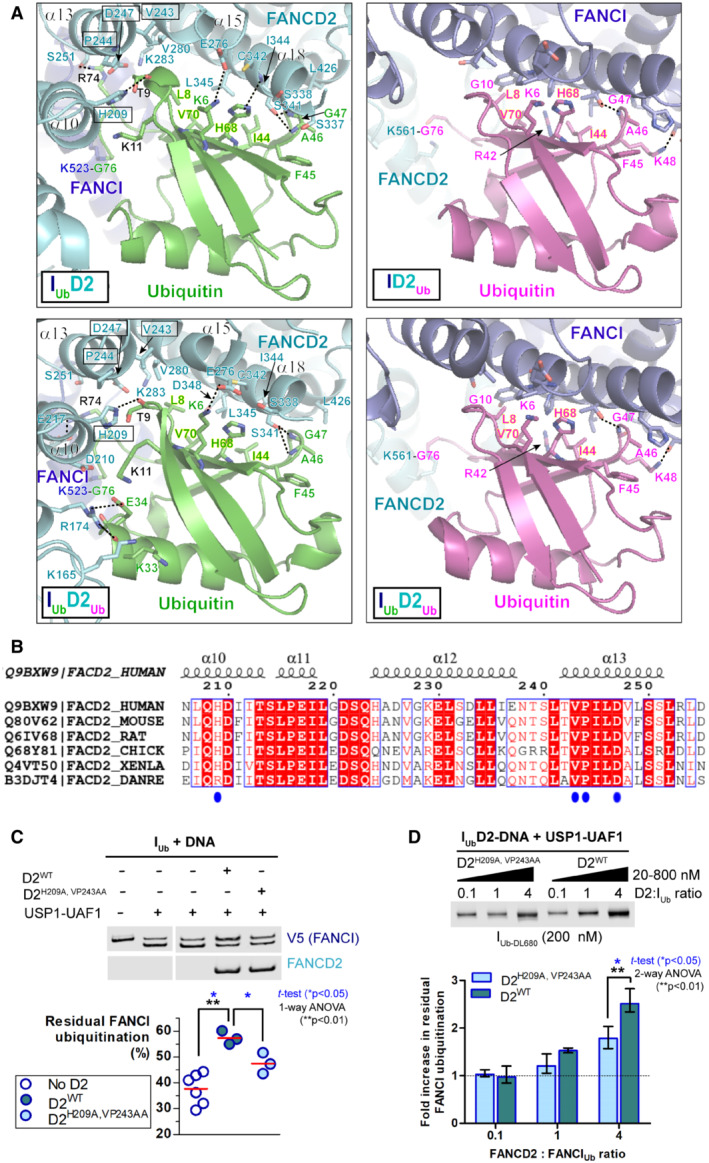
- A
Interactions of FANCI and FANCD2 with the ubiquitin conjugated to the other ID2 subunit, in DNA‐bound IUbD2, IUbD2Ub (PDB:
6VAE ), ID2Ub (PDB:6VAF ) and IUbD2Ub (PDB:6VAE ) structures (Wang et al, 2020). Dotted straight lines indicate hydrogen bonding. Both FANCD2 and FANCI interact with ubiquitin's hydrophobic I44 patch (residues L8, I44, H68 and V70; all labelled in highlighted‐yellow font) and additionally with residues F45 to G47 of ubiquitin. However, the ubiquitin conjugated to FANCI has a more extensive interface with FANCD2. This extended interface is formed by interactions of FANCD2 α10 ‐ α13 helices (predominant interacting residues highlighted in boxes) with residues R74, T9 and K11 of ubiquitin (shown in black font). The ubiquitin‐FANCD2 interface may be further extended via interactions between residues K33 and E34 of ubiquitin with K165 and R174 of FANCD2, as shown in IUbD2Ub‐DNA structure. For direct comparison of corresponding interactions, the same orientation for all ubiquitins (both FANCI‐conjugated and FANCD2‐conjugated) was achieved by aligning: ID2Ub‐DNA and IUbD2Ub‐DNA structures to the ubiquitin of IUbD2‐DNA structure, and subsequently, the IUbD2Ub‐DNA structure to the ubiquitin of ID2Ub‐DNA structure as well. - B
Clustal O multiple sequence alignment of human, mouse, rat, chicken, frog and zebrafish FANCD2 amino‐acid sequences, focused on a region encompassing α11‐13 helices of FANCD2 in human IUbD2‐DNA structure (helical regions shown on top). Identical residues among various species are highlighted red, whereas residues in positions displaying 83% similarity/identity are shown in red font. Residues of FANCD2 interacting with FANCI's ubiquitin in both IUbD2‐DNA and IUbD2Ub‐DNA structures (extended FANCD2‐ubiquitin interface) are indicated with blue circles.
- C, D
Alanine mutagenesis of key FANCD2's residues (H209, V243 and P244) participating in the extended FANCD2‐ubiquitin interface (IUbD2‐DNA and IUbD2Ub‐DNA structures), results in enhanced FANCI deubiquitination by USP1‐UAF1. (C) FANCI deubiquitination by USP1‐UAF1 (incubation with 50 nM USP1‐UAF1 for 30 min at room temperature) in the presence of DNA and, in the absence or presence of wild‐type (D2WT) or mutant (D2H209A,VP243AA) FANCD2. Replicate residual ubiquitination values (three technical replicates) and statistically significant changes (t‐test and one‐way ANOVA test with Bonferroni correction) are shown. (D) FANCI was ubiquitinated with DyLight‐680 labelled ubiquitin (IUb‐DL680) and its deubiquitination by USP1‐UAF1 (incubation with 100 nM USP1‐UAF1 for 20 min at room temperature), was assessed at increasing concentrations (20, 200 or 200 nM) of, either wild‐type (D2WT), or mutant (D2H209A,VP243AA) FANCD2. Experiment was repeated three times (technical replicates), and the fold change in residual FANCI ubiquitination was determined by normalisation to the mean intensity of IUb‐DL680 at 20 nM FANCD2 (average from six values, for both D2WT and D2H209A,VP243AA). Mean (with range) fold‐increase values and statistically significant changes upon FANCD2 mutation for each FANCD2 concentration (t‐test and two‐way ANOVA test with Bonferroni correction) are shown.
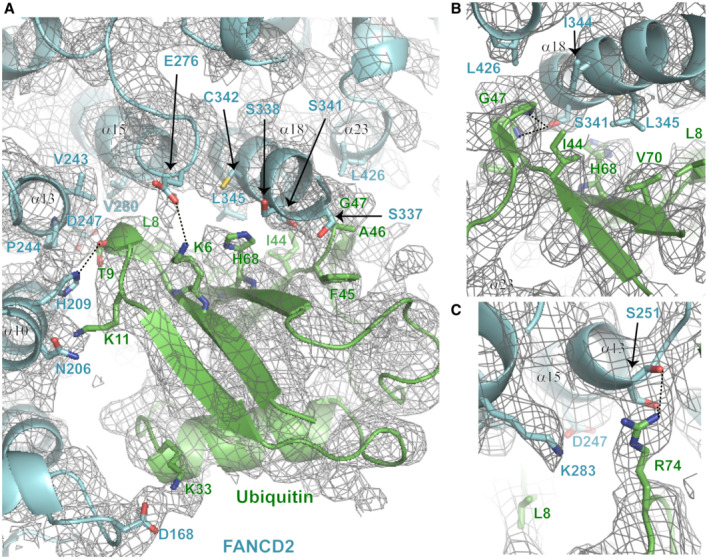
The ubiquitin conjugated to FANCI interacts with several residues located in helices α10, α13, α15, α18 and α23 of FANCD2 in IUbD2‐DNA structure. Assessment of the density map (Phenix auto‐sharpened IUbD2‐DNA map) and of the nature and distance between interacting residues, indicate that the ubiquitin of FANCI predominantly interacts with residues located on helices α10 (H209) and α13 (V243 and P244), and on helix α18 (S337, S338, S341, C342) of FANCD2. Interacting residues are indicated and illustrated as sticks. D168 and N206 of FANCD2 (also shown and illustrated as sticks) are positioned in areas corresponding to overlapping density between ubiquitin and FANCD2, but are not sufficiently close to K33 and K11 of ubiquitin, respectively, for a high confidence interaction.
Ubiquitin‐FANCD2 interface from a view centred on residues G47, I44, H68, V70 and L8 of ubiquitin.
Ubiquitin‐FANCD2 interface from a view centred on residues K283 and S251 of FANCD2.
Similar articles
-
Mechanism, specificity, and function of FANCD2-FANCI ubiquitination and deubiquitination.FEBS J. 2022 Aug;289(16):4811-4829. doi: 10.1111/febs.16077. Epub 2021 Jun 29. FEBS J. 2022. PMID: 34137174 Review.
-
Differential functions of FANCI and FANCD2 ubiquitination stabilize ID2 complex on DNA.EMBO Rep. 2020 Jul 3;21(7):e50133. doi: 10.15252/embr.202050133. Epub 2020 Jun 8. EMBO Rep. 2020. PMID: 32510829 Free PMC article.
-
The FANCD2-FANCI complex is recruited to DNA interstrand crosslinks before monoubiquitination of FANCD2.Nat Commun. 2016 Jul 13;7:12124. doi: 10.1038/ncomms12124. Nat Commun. 2016. PMID: 27405460 Free PMC article.
-
Mechanism of Ubiquitination and Deubiquitination in the Fanconi Anemia Pathway.Mol Cell. 2017 Jan 19;65(2):247-259. doi: 10.1016/j.molcel.2016.11.005. Epub 2016 Dec 13. Mol Cell. 2017. PMID: 27986371
-
The Fanconi anemia ID2 complex: dueling saxes at the crossroads.Cell Cycle. 2014;13(19):2999-3015. doi: 10.4161/15384101.2014.956475. Cell Cycle. 2014. PMID: 25486561 Free PMC article. Review.
Cited by
-
Repair and tolerance of DNA damage at the replication fork: A structural perspective.Curr Opin Struct Biol. 2023 Aug;81:102618. doi: 10.1016/j.sbi.2023.102618. Epub 2023 Jun 1. Curr Opin Struct Biol. 2023. PMID: 37269798 Free PMC article. Review.
-
DoUBLing up: ubiquitin and ubiquitin-like proteases in genome stability.Biochem J. 2024 Apr 10;481(7):515-545. doi: 10.1042/BCJ20230284. Biochem J. 2024. PMID: 38572758 Free PMC article. Review.
-
STIM1 translocation to the nucleus protects cells from DNA damage.Nucleic Acids Res. 2024 Mar 21;52(5):2389-2415. doi: 10.1093/nar/gkae001. Nucleic Acids Res. 2024. PMID: 38224453 Free PMC article.
-
Phosphorylation by ATR triggers FANCD2 chromatin loading and activates the Fanconi anemia pathway.Cell Rep. 2023 Jul 25;42(7):112721. doi: 10.1016/j.celrep.2023.112721. Epub 2023 Jun 30. Cell Rep. 2023. PMID: 37392383 Free PMC article.
References
-
- Chaugule VK, Arkinson C, Toth R, Walden H (2019) Enzymatic preparation of monoubiquitinated FANCD2 and FANCI proteins. Methods Enzymol 618: 1–30 - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
