

Sexually dimorphic architecture and function of a mechanosensory circuit in C. elegans
- PMID: 36369281
- PMCID: PMC9652301
- DOI: 10.1038/s41467-022-34661-3
Sexually dimorphic architecture and function of a mechanosensory circuit in C. elegans
Abstract
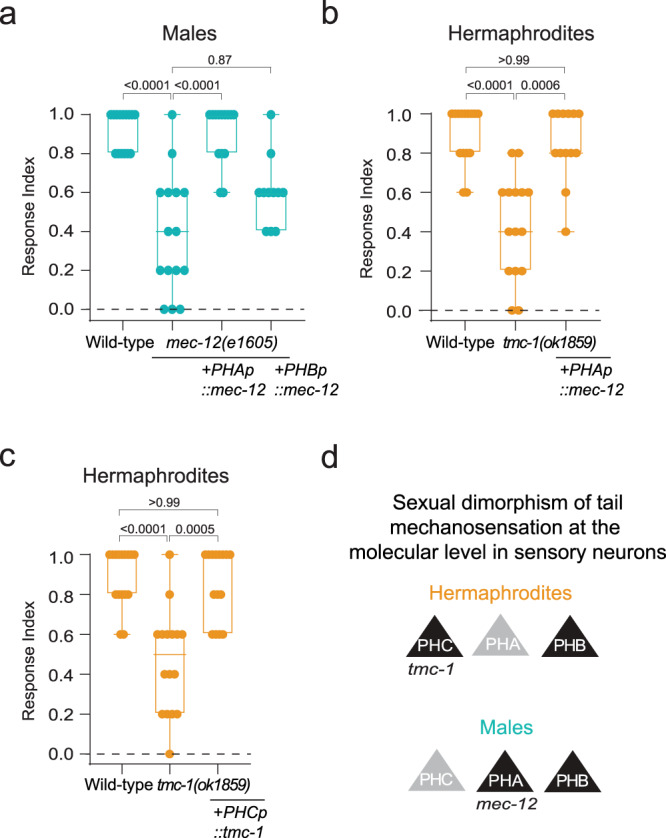
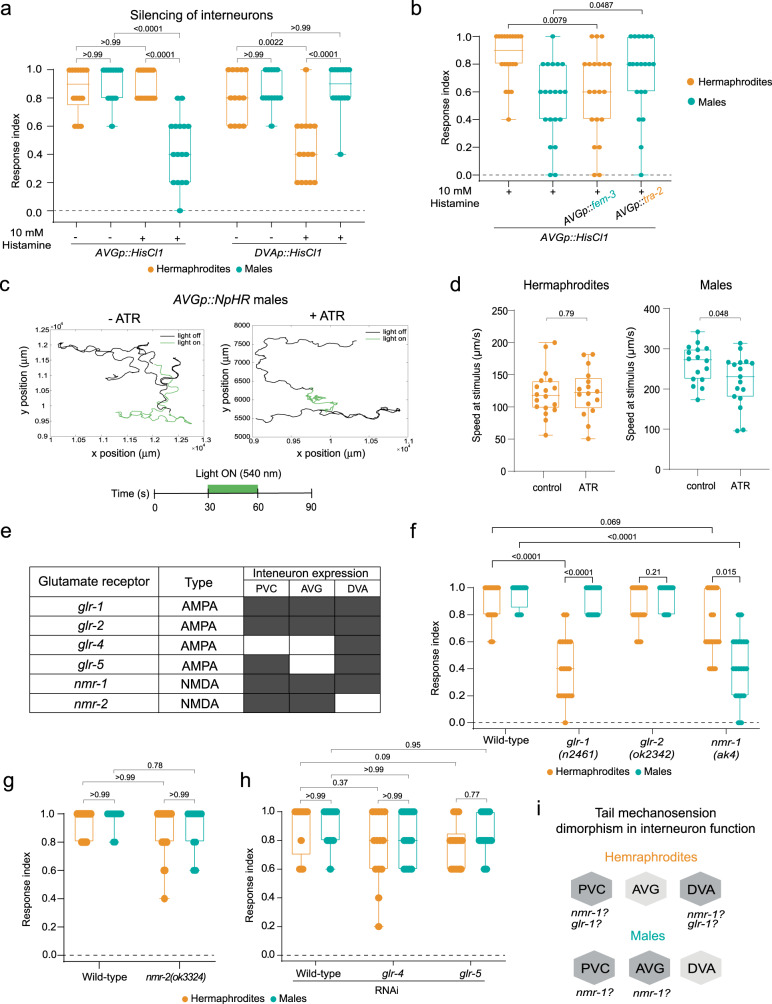
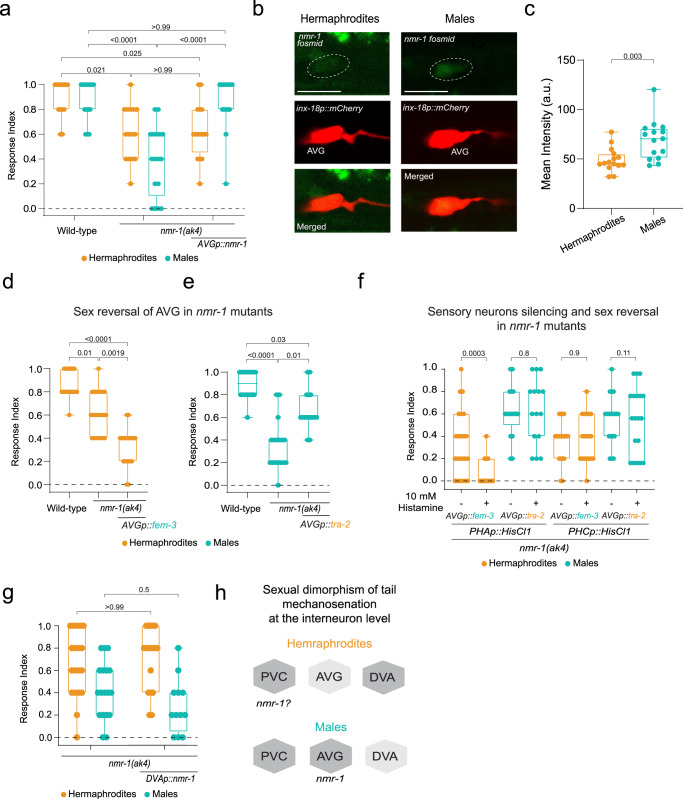
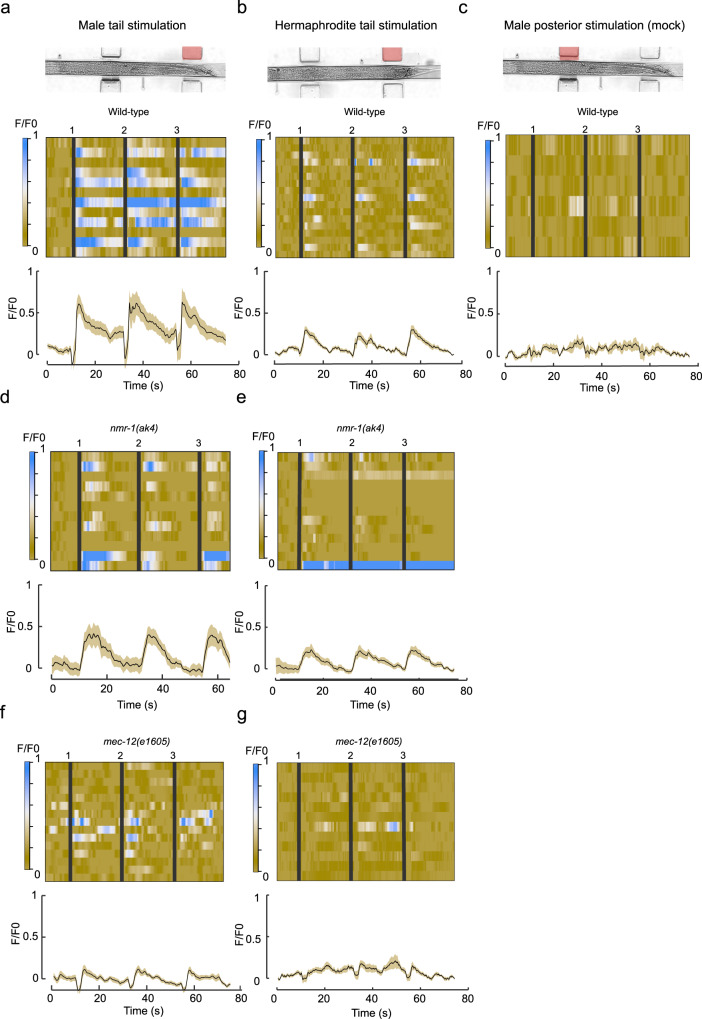
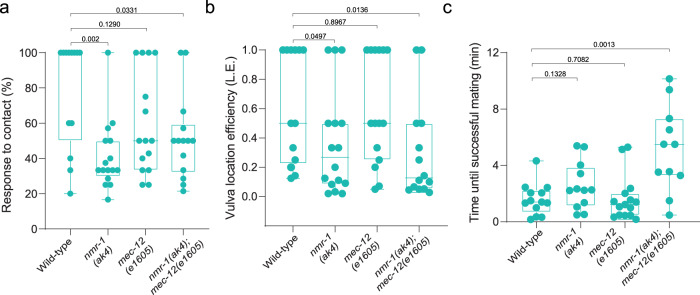
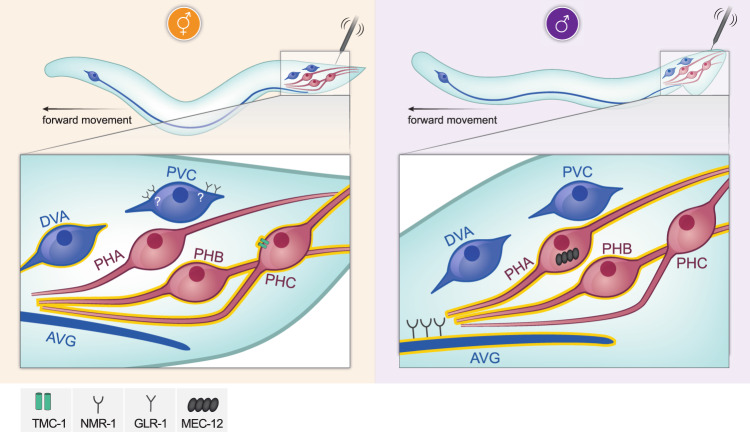
How sensory perception is processed by the two sexes of an organism is still only partially understood. Despite some evidence for sexual dimorphism in auditory and olfactory perception, whether touch is sensed in a dimorphic manner has not been addressed. Here we find that the neuronal circuit for tail mechanosensation in C. elegans is wired differently in the two sexes and employs a different combination of sex-shared sensory neurons and interneurons in each sex. Reverse genetic screens uncovered cell- and sex-specific functions of the alpha-tubulin mec-12 and the sodium channel tmc-1 in sensory neurons, and of the glutamate receptors nmr-1 and glr-1 in interneurons, revealing the underlying molecular mechanisms that mediate tail mechanosensation. Moreover, we show that only in males, the sex-shared interneuron AVG is strongly activated by tail mechanical stimulation, and accordingly is crucial for their behavioral response. Importantly, sex reversal experiments demonstrate that the sexual identity of AVG determines both the behavioral output of the mechanosensory response and the molecular pathways controlling it. Our results present extensive sexual dimorphism in a mechanosensory circuit at both the cellular and molecular levels.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
The Snail transcription factor CES-1 regulates glutamatergic behavior in C. elegans.PLoS One. 2021 Feb 2;16(2):e0245587. doi: 10.1371/journal.pone.0245587. eCollection 2021. PLoS One. 2021. PMID: 33529210 Free PMC article.
-
A Single-Neuron Chemosensory Switch Determines the Valence of a Sexually Dimorphic Sensory Behavior.Curr Biol. 2018 Mar 19;28(6):902-914.e5. doi: 10.1016/j.cub.2018.02.029. Epub 2018 Mar 8. Curr Biol. 2018. PMID: 29526590 Free PMC article.
-
EGL-4/PKG regulates the role of an interneuron in a chemotaxis circuit of C. elegans through mediating integration of sensory signals.Genes Cells. 2021 Jun;26(6):411-425. doi: 10.1111/gtc.12849. Epub 2021 Apr 16. Genes Cells. 2021. PMID: 33817914
-
Mechanosensation circuitry in Caenorhabditis elegans: A focus on gentle touch.Peptides. 2015 Jun;68:164-74. doi: 10.1016/j.peptides.2014.12.004. Epub 2014 Dec 25. Peptides. 2015. PMID: 25543196 Review.
-
Sexual modulation of sex-shared neurons and circuits in Caenorhabditis elegans.J Neurosci Res. 2017 Jan 2;95(1-2):527-538. doi: 10.1002/jnr.23912. J Neurosci Res. 2017. PMID: 27870393 Free PMC article. Review.
Cited by
-
Sex-specific developmental gene expression atlas unveils dimorphic gene networks in C. elegans.Nat Commun. 2024 May 20;15(1):4273. doi: 10.1038/s41467-024-48369-z. Nat Commun. 2024. PMID: 38769103 Free PMC article.
-
Biophysical modeling of the whole-cell dynamics of C. elegans motor and interneurons families.PLoS One. 2024 Mar 29;19(3):e0298105. doi: 10.1371/journal.pone.0298105. eCollection 2024. PLoS One. 2024. PMID: 38551921 Free PMC article.
-
Neural Circuit Remodeling: Mechanistic Insights from Invertebrates.J Dev Biol. 2024 Oct 11;12(4):27. doi: 10.3390/jdb12040027. J Dev Biol. 2024. PMID: 39449319 Free PMC article. Review.
-
The synaptic basis for sexual dimorphism in the invertebrate nervous system.Curr Opin Neurobiol. 2023 Oct;82:102757. doi: 10.1016/j.conb.2023.102757. Epub 2023 Aug 10. Curr Opin Neurobiol. 2023. PMID: 37572555 Free PMC article. Review.
-
Automated dual olfactory device for studying head/tail chemosensation in Caenorhabditis elegans.APL Bioeng. 2024 Apr 18;8(2):026104. doi: 10.1063/5.0187441. eCollection 2024 Jun. APL Bioeng. 2024. PMID: 38655543 Free PMC article.
References
-
- Shen J-X, et al. Ultrasonic frogs show extraordinary sex differences in auditory frequency sensitivity. Nat. Commun. 2011;2:342. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
