

Disrupted-in-schizophrenia-1 is required for normal pyramidal cell-interneuron communication and assembly dynamics in the prefrontal cortex
- PMID: 36239988
- PMCID: PMC9566853
- DOI: 10.7554/eLife.79471
Disrupted-in-schizophrenia-1 is required for normal pyramidal cell-interneuron communication and assembly dynamics in the prefrontal cortex
Abstract
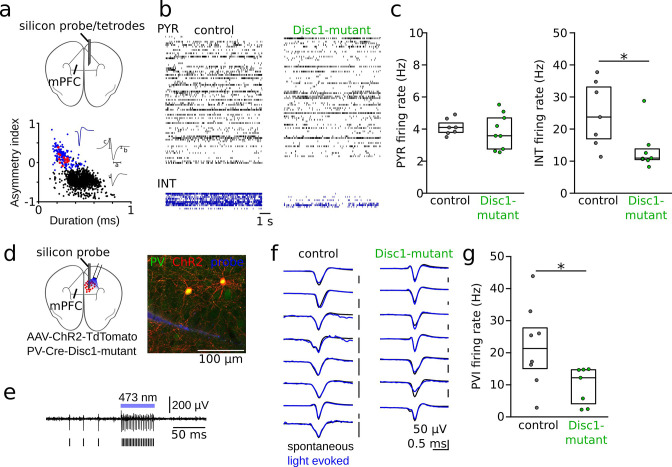
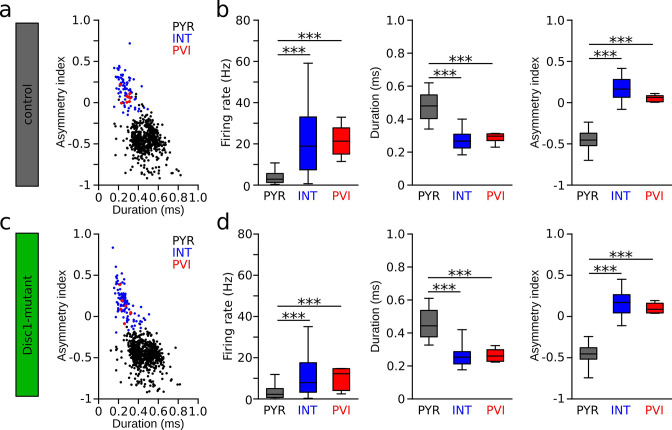
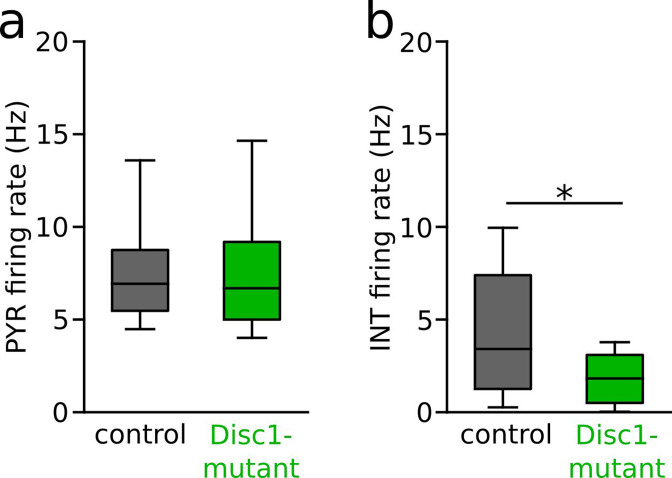
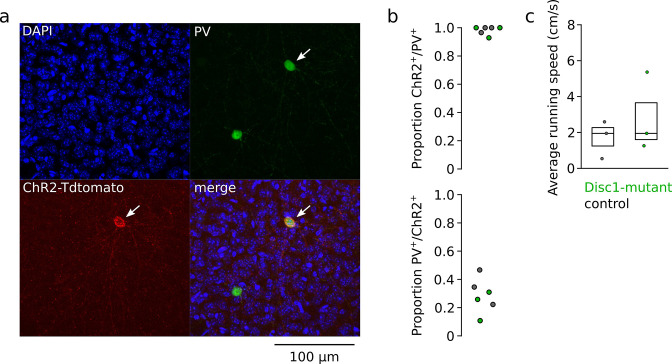
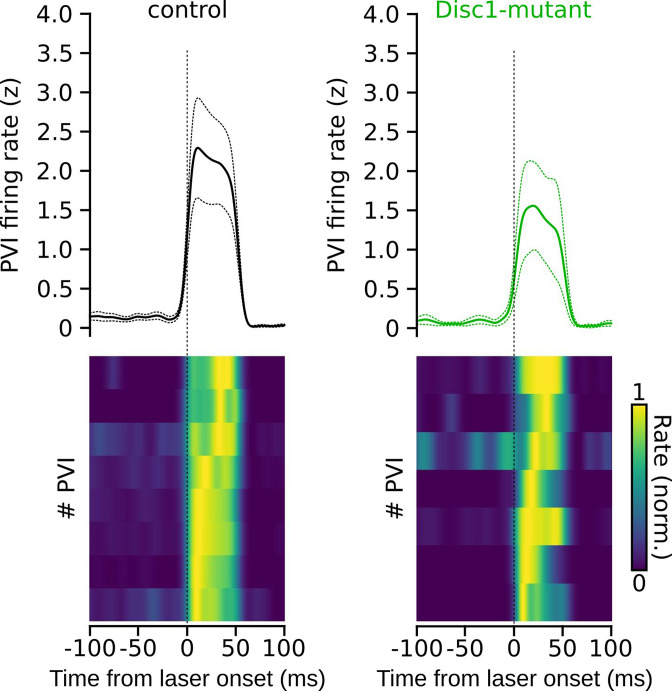
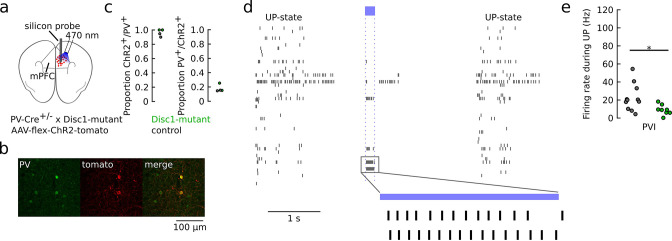
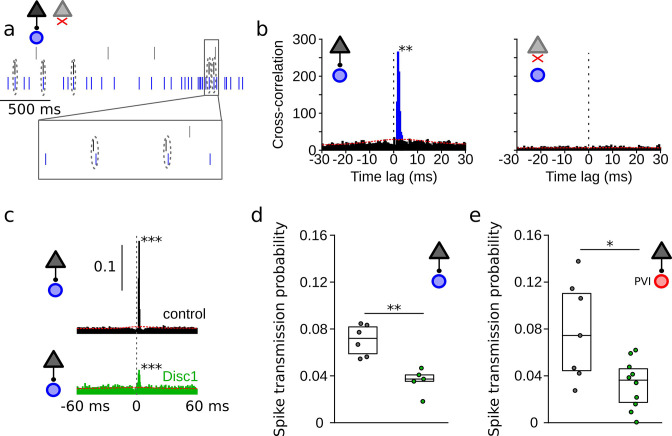
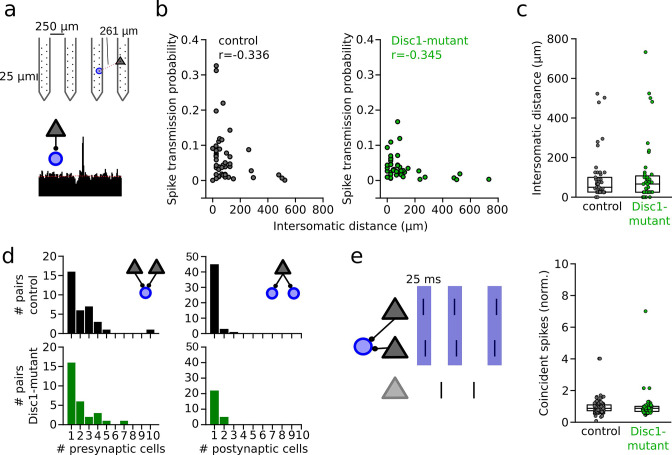
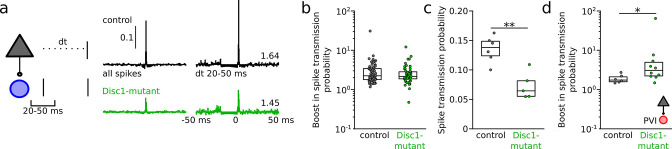
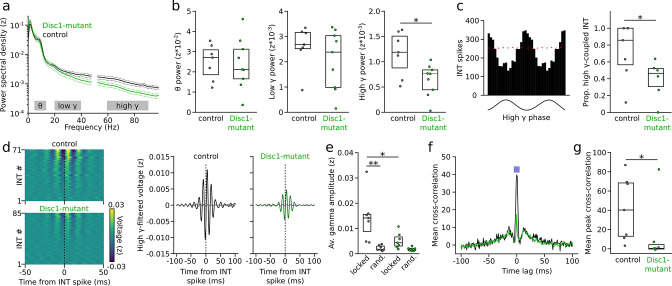
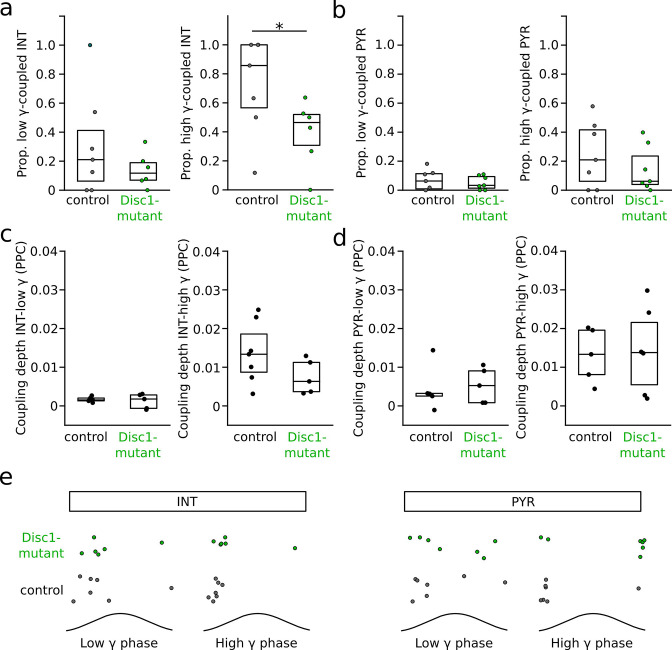
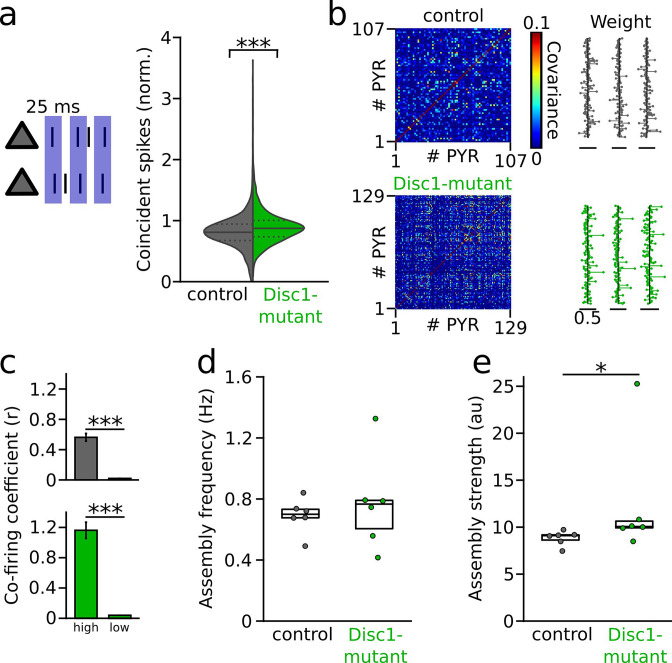
We interrogated prefrontal circuit function in mice lacking Disrupted-in-schizophrenia-1 (Disc1-mutant mice), a risk factor for psychiatric disorders. Single-unit recordings in awake mice revealed reduced average firing rates of fast-spiking interneurons (INTs), including optogenetically identified parvalbumin-positive cells, and a lower proportion of INTs phase-coupled to ongoing gamma oscillations. Moreover, we observed decreased spike transmission efficacy at local pyramidal cell (PYR)-INT connections in vivo, suggesting a reduced excitatory effect of local glutamatergic inputs as a potential mechanism of lower INT rates. On the network level, impaired INT function resulted in altered activation of PYR assemblies: While assembly activations defined as coactivations within 25 ms were observed equally often, the expression strength of individual assembly patterns was significantly higher in Disc1-mutant mice. Our data, thus, reveal a role of Disc1 in shaping the properties of prefrontal assembly patterns by setting INT responsiveness to glutamatergic drive.
Keywords: DISC1; interneuron; mouse; neuronal assembly; neuroscience; prefrontal cortex; schizophrenia; synaptic transmission.
© 2022, Sauer and Bartos.
Conflict of interest statement
JS, MB No competing interests declared
Figures
Similar articles
-
Parvalbumin Interneuron Dysfunction in a Thalamo-Prefrontal Cortical Circuit in Disc1 Locus Impairment Mice.eNeuro. 2020 Mar 3;7(2):ENEURO.0496-19.2020. doi: 10.1523/ENEURO.0496-19.2020. Print 2020 Mar/Apr. eNeuro. 2020. PMID: 32029441 Free PMC article.
-
Abnormal interneuron development in disrupted-in-schizophrenia-1 L100P mutant mice.Mol Brain. 2013 Apr 30;6:20. doi: 10.1186/1756-6606-6-20. Mol Brain. 2013. PMID: 23631734 Free PMC article.
-
Impaired fast-spiking interneuron function in a genetic mouse model of depression.Elife. 2015 Mar 3;4:e04979. doi: 10.7554/eLife.04979. Elife. 2015. PMID: 25735038 Free PMC article.
-
Cortical parvalbumin interneurons and cognitive dysfunction in schizophrenia.Trends Neurosci. 2012 Jan;35(1):57-67. doi: 10.1016/j.tins.2011.10.004. Epub 2011 Dec 6. Trends Neurosci. 2012. PMID: 22154068 Free PMC article. Review.
-
Prefrontal Interneurons: Populations, Pathways, and Plasticity Supporting Typical and Disordered Cognition in Rodent Models.J Neurosci. 2022 Nov 9;42(45):8468-8476. doi: 10.1523/JNEUROSCI.1136-22.2022. J Neurosci. 2022. PMID: 36351822 Free PMC article. Review.
Cited by
-
Neuronal tuning to threat exposure remains stable in the mouse prefrontal cortex over multiple days.PLoS Biol. 2024 Jan 11;22(1):e3002475. doi: 10.1371/journal.pbio.3002475. eCollection 2024 Jan. PLoS Biol. 2024. PMID: 38206890 Free PMC article.
-
Inhibitory circuits in fear memory and fear-related disorders.Front Neural Circuits. 2023 Mar 23;17:1122314. doi: 10.3389/fncir.2023.1122314. eCollection 2023. Front Neural Circuits. 2023. PMID: 37035504 Free PMC article. Review.
-
Large-scale coupling of prefrontal activity patterns as a mechanism for cognitive control in health and disease: evidence from rodent models.Front Neural Circuits. 2024 Apr 4;18:1286111. doi: 10.3389/fncir.2024.1286111. eCollection 2024. Front Neural Circuits. 2024. PMID: 38638163 Free PMC article. Review.
-
Dysregulated Signaling at Postsynaptic Density: A Systematic Review and Translational Appraisal for the Pathophysiology, Clinics, and Antipsychotics' Treatment of Schizophrenia.Cells. 2023 Feb 10;12(4):574. doi: 10.3390/cells12040574. Cells. 2023. PMID: 36831241 Free PMC article. Review.
-
Behavioral State-Dependent Modulation of Prefrontal Cortex Activity by Respiration.J Neurosci. 2023 Jun 28;43(26):4795-4807. doi: 10.1523/JNEUROSCI.2075-22.2023. Epub 2023 Jun 5. J Neurosci. 2023. PMID: 37277176 Free PMC article.
References
-
- Akbarian S, Kim JJ, Potkin SG, Hagman JO, Tafazzoli A, Bunney WE, Jones EG. Gene expression for glutamic acid decarboxylase is reduced without loss of neurons in prefrontal cortex of schizophrenics. Archives of General Psychiatry. 1995;52:258–266. doi: 10.1001/archpsyc.1995.03950160008002. - DOI - PubMed
-
- Blackwood DH, Fordyce A, Walker MT, St Clair DM, Porteous DJ, Muir WJ. Schizophrenia and affective disorders -- cosegregation with a translocation at chromosome 1q42 that directly disrupts brain-expressed genes: clinical and p300 findings in a family. American Journal of Human Genetics. 2001;69:428–433. doi: 10.1086/321969. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
